Open Access
Research Article
Max Screen
ISSN: 2348-9790
Copyright: © 2023 Yang Yingkui. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
The study investigated the effects of supplementing the same level of concentrate in different seasons on the growth performance, rumen fermentation, serum biochemical index, and rumen microflora changes in yaks, to provide guidance on accurate feeding of yaks in cold and warm seasons. 12 healthy yaks weighing (124.21±15.64) kg were selected for the experiment, with a warm season group from August to October and a Cold season supplemental feeding group from October to December. The results showed that the mean daily gain was significantly higher in the warm season grazing group than in the cold season grazing group (p< 0.05). Rumen fermentation, serum biochemical index, and rumen microflora changes also varied significantly between the two groups (p< 0.05). The study suggests that supplementing the concentrate above 2.5kg/d per head in the cold season can promote the growth and health of grazing yaks.
Keywords: Same Supplementation Level; Seasonal Variation; Growth Performance; Rumen Fermentation; Serum Biochemical Indexes; Rumen Microbiota Structure
Abbreviations: CP: Crude protein; EE: Ether Extract; NDF: Neutral detergent fiber; ADF: Acid detergent fiber; NH3 -N: Ammoniacal nitrogen; MCP: Ruminal microbial protein; WGYS: Warm season supplementary feeding group; GYS: Cold season supplementary feeding group
In the Tibetan Plateau region, there is a unique climatic environment. The average altitude is over 3,000 meters, and the average annual temperature is below 5°C in most areas, except for the valleys in the southeastern part of the plateau, where it is higher. In other regions, the seasons are divided into "spring, summer, autumn, and winter", while the average temperature of the Tibetan Plateau is lower than 0°C throughout the year, except for June to September, when the temperature is higher [1]. Therefore, the seasonal distinction on the Qinghai-Tibet Plateau is based on the "cold season" and "warm season". Both cold and warm seasons are subject to sudden changes in temperature, especially in these years when extreme weather is common [2]. In the Tibetan plateau, yak is the main livestock, and the breeding mode is mostly a combination of grazing and supplemental feeding. However, the lack of supplemental feeding concentrate resources and high transportation costs in pastoral areas cause many herders not to supplement their yaks and Tibetan sheep.
Due to the unique climate, many yaks and Tibetan sheep will lose fat in the cold season without supplemental feeding, which seriously affects the income of herders. Whether it is a combination of grazing and supplemental feeding, climate change, or pure grazing patterns there will be changes in herd and diet changes which will cause stress to the yaks. The process from the time the stress is caused to the time the stress disappears is called the adaptation period, which lasts about 15 days. It has been shown that different energy levels can affect the rumen microbial structure of ruminants, thus affecting the adaptation period to feed changes [3].
The duration of the acclimation period of Tibetan plateau yaks to different feeds may vary depending on various factors such as individual differences, feed composition and feeding methods. In general, the composition of rumen microorganisms and digestive enzymes needs to be changed in order to adapt to the digestion and absorption of different feeds when Tibetan plateau yaks suffer from starvation or food changes. This adaptation period usually takes 3 to 4 weeks, but in special cases it may take longer. In addition, if the feed changeover process is too fast, the yak's digestive system may not be able to adapt, leading to problems such as indigestion, diarrhea and low appetite. Therefore, you should gradually increase the new feed and reduce the proportion of the old feed during the changeover process and make sure the transition between the old and new feeds is smooth. At the same time, the appetite and fecal condition as well as weight change and other indicators of yaks should be closely observed to adjust feed composition and feeding amount in time to ensure healthy growth of yaks.
For ruminants, the warm season is the main season for forage growth, while the cool season is relatively dry and lacking in forage. During the warm season temperatures rise and forage begins to grow, which is very beneficial to ruminants. Forage in season is nutrient-rich and fresh, which positively affects the health status, growth and development, and reproduction of ruminants. The long hours of sunshine, combined with abundant vegetation and a richer and more varied forage than in the cool season, also contribute to the health of ruminants. During the cold season, as the weather gradually becomes colder, the amount of grass decreases and attention needs to be paid to supplementing feed to maintain normal growth and development of ruminants. The cold season is colder and the amount of grass becomes less, so ruminants need to be managed with proper nutrition. In addition, attention needs to be paid to issues such as keeping warm and preventing infections and diseases to ensure that ruminants can grow well and healthy even during the cold season. In conclusion, different seasons will have an impact on the health of ruminants, and reasonable feeding management and adjustment of feed combinations are the keys to ensure the health of ruminants.
In a previous study of supplemental feeding for warm-season grazing yaks, our team found that supplemental feeding of 2.5 kg per head per day improved growth performance and rumen microecological stability compared with other supplemental feeds. On this basis, we supplemented each cow with 2.5kg of feed every day to explore the effects of seasonal changes on yak growth performance, rumen fermentation, serum biochemical parameters, and rumen microbial structure.
The trial was conducted from August 2019 to February 2020 at Laozashi Ranch in Guinan, Qinghai Province.
In this experiment, 12 yaks in good condition, free from disease and weighing (124.21±15.64) kg were selected. The warm season group was two months from August to October, and the buffer period was from October to December, when the yaks were fed 2.5 kg of concentrate without counting the data. The trial period was 70 days, with a pre-test period of 10 days and a positive test period of 60 days.
All test cattle were grazed in the same pasture, starting grazing at 08:00 each day and collecting cattle at 18:00 each day. Supplementary feeding was done in two parts before grazing and after grazing, and all test yaks had unrestricted access to water.
Supplemental feeding concentrate was prepared with reference to beef cattle feeding standards (NY/T815–2004) and combined with grazing yak feed intake. The nutritional level, concentrate composition and concentrate nutrition level of forage grass are shown in Table 1.
1) The premix provided the following per kg of diets:Cu 10mg,Fe 65mg,Mn 30mg,Zn 25mg,I 0.5mg,Se 0.1mg,Co 0.1mg,VA
1.2×10-3g,VD 1.5×10-4g,VE 1.2×10-5g。
2) ME was a calculated value[10],while the others were measured values。
CP: Crude protein;EE: Ether Extract;NDF: Neutral detergent fiber;ADF: Acid detergent fiber.
On the 60th day of the formal test, 150mL of rumen fluid was collected by gastric tube sampler (GCYQ-1-A, Wuhan Clebo Equipment Co., Ltd.) before morning feeding, and the pH value was determined immediately after filtering with 4 layers of gauze, and the remaining rumen juice samples were aliquoted into 15mL centrifuge tubes, immediately stored at -80℃, and transported to the laboratory.
Before the start of the trial period and before the end of the morning feeding, the average initial weight, average final weight, total weight gain and average daily weight gain were calculated for 2 consecutive days
Total weight gain = average final weight - average initial weight [4]
Average daily gain = total gain ÷ 60
Average net profit = total weight gain× live cattle price - daily concentrate supplementation × price × 60
At the end of the trial, 12 yaks per group, 10 mL of jugular venous blood was collected, left to stand for 10min at room temperature, centrifuged at 3000 r/min for 10min, and stored at -20°C in the upper serum. The concentration of blood glucose (GLU) [5], total protein (TP), albumin (ALB), globulin (GLO), total cholesterol (TCH), triglycerides (TG), urea nitrogen (BUN) and the activity of alanine aminotransferase (ALT) and glutamate aminotransferase (AST) were commissioned by Qinghai Provincial People's Hospital.
pH: The pH of rumen fluid was determined by using a benchtop pH meter (HANNAHI221, Beijing Jingmeirui Technology Co., Ltd.), and the acidity meter was corrected with the corresponding standard solution before the measurement; Referring to the improved colorimetric method of Feng Zongci et al[6]. To determine the concentration of rumen liquid ammonia nitrogen (NH3-N), the instrument is ultraviolet-visible spectrophotometer (TU-1810, Beijing General Instrument Co., Ltd.), preheated for 30min, the optical density (OD) value of the solution is determined at a wavelength of 625nm, and the NH3-N concentration of the sample is determined by standard curve; Microprotein (MCP) concentration was determined colorimetrically at 595 nm by Coomassie Brilliant Blue Method [7] (A045-2, kit purchased in Nanjing Institute of Bioengineering); The concentration of volatile fatty acids (VFA) was determined using the Shimadzu GC-2014 gas chromatograph. Among them, the VFA concentration determination conditions were: hydrogen flame ion detector (FID, Shimadzu Co., Ltd.), and the column was a capillary column (FFAP, 30.00m×0.32mm×0.50μm); The heating conditions were as follows: the initial 60 °C, the temperature was raised to 120 °C at 10 °C·min-1, and the temperature was retained for 2min, and the temperature was raised to 180 °C at 15 °C·min-1, retained for 5min, and the temperature of the vaporization chamber was 250 °C. FID temperature 250°C; The injection amount is 1μL, the carrier gas is high-purity nitrogen (99.99%), the pressure is 0.7MPa, the hydrogen pressure is 0.4MPa, the air pressure is 0.4MPa, the capillary column pressure is 0.6~0.8MPa, and the split ratio is 40:1.
Each group randomly selected the rumen juice samples of 4 cows were thawed on ice and mixed thoroughly, and the genomic DNA of rumen samples was extracted by referring to the CTAB method of Fan J et al [8], the purity and concentration of DNA were detected by 1.0% agarose gel electrophoresis, an appropriate amount of sample was taken in a centrifuge tube, and the sample was diluted with sterile water to 1ng·μL-1[9]. According to the V3~V4 region of the bacterial 16SrDNA gene, specific primers with Barcode were synthesized. Universal primer sequence 515F: 5'-GTGCCAGCMGCCGCGG-3', 806R:5'-GGACTACHVGGG TWTCTAAT-3'[10]. PCR uses a 25 μL amplification system; DNA template 5ng, 5 μmol· 1μL of L-1 upstream and downstream primers, 3μL of bovine serum albumin (BSA) of 2ng·μL-1, 2×Taq Plus Master Mix 12.5 μL, double distilled water (ddH2O) added to 25 μL; PCR amplification conditions: predenaturation at 94°C for 5min; 94 °C denaturation for 30s, 50 °C annealing for 30s, 72 °C extension for 60s, a total of 30 cycles; Extend at 72 °C for 7 min. The PCR amplification product was detected by 1.0% agarose gel electrophoresis, purified and recovered by QIAquickPCR purification kit (Qiagen, Germany), and the DNA sample that met the requirements was sent to Beijing Avison Gene Technology Co., Ltd. for sequencing, and the sequencing platform was Mixed PE300.
The raw data obtained by the sequencing platform were obtained using Trimmomatic (Version 0. 36). The chimeras were then removed by vsearch software and species databases to obtain valid sequences. OTUs were generated by screening sequences with similarity above 97%, and then OTU clustering, species taxonomy, diversity index, and community structure were statistically analyzed.
The test data were initially collated using Excel 2021 and t-tested using SPSS 22.0. p< 0.05 indicates a significant difference and p< 0.01 indicates a highly significant difference. Results are shown as mean ± standard deviation.
As shown in Table 2, the total weight gain and average daily gain of the warm season feeding group were significantly higher than that of the cold season feeding group (p< 0.05), and the total weight gain and average daily weight gain of the warm season feeding group increased by 82.39% and the average daily weight gain increased by 82.61% compared with the cold season feeding group. The warm season feeding group gained 0.38kg / head per day compared with the cold season supplementary feeding group. According to the price of yak live cattle of 31 yuan/kg and the price of concentrate of 3.5 yuan/kg, the average net profit per head increased by 214.84% compared with the cold season feeding group during the trial period. That is, the net profit increased by 700.29 yuan per head.
WGYS: Warm season supplementary feeding group
GYS: Cold season supplementary feeding group
This is shown in Table 3,In this experiment, the same level of supplementation showed that the pH of the warm season feeding group was higher than that of the cold season feeding group, but the difference was not significant (p>0.05), and the ammonia nitrogen cold season feeding group was significantly higher than that of the warm season feeding group (p< 0.01). The microbial protein warm season feeding group was higher than that in the cold season feeding group, and the difference was not significant (p>0.05). The cold season supplementation group of volatile fatty acids acetic acid was significantly higher than that of the warm season feeding group (p< 0.05), the propionic acid warm season feeding group was significantly higher than the cold season feeding group (p < 0.01), the isobutyric acid warm season feeding group was significantly higher than the cold season feeding group (p< 0.01), the butyric acid warm season feeding group was higher than the cold season feeding group but the difference was not significant (p>0.05), and the isovaleric acid and valeric acid warm season feeding group were significantly higher than the cold season feeding group (p< 0.01). The acetic acid/ propionic acid ratio was highly significant higher in the cold season supplementation group than in the warm season supplementation group (p< 0.01).
This is shown in Table 4, In this experiment, the effect of the same level of supplementation on serum biochemical indexes of grazing yaks in different seasons was that the glucose cold season feeding group was higher than that of the warm season feeding group, the cholesterol warm season feeding group was higher than the cold season feeding group, the globulin warm season feeding group was higher than the cold season feeding group, the gluten aminotransferase warm season feeding group was higher than the cold season feeding group, and the alkaline phosphatase cold season feeding group was higher than the warm season feeding group, but the difference was not significant (p>0.05). The total protein warm season feeding group was significantly higher than that of the cold season feeding group (p< 0.05), and the albumin warm season feeding group was significantly higher than that of the cold season feeding group (p< 0.05). The urea nitrogen cold season feeding group was significantly higher than that in the warm season feeding group (p< 0.01), and the alanine aminotransferase warm season feeding group was significantly higher than that in the cold season feeding group (p< 0.01)
At the phylum level, a total of 21 phylums were detected. Table 6, Firmicutes and Bacteroides were the dominant phyla, and Firmicutes accounted for 64.61% and 60.69% in the warm season and cold season feeding groups, respectively. Bacteroides accounted for 25.93% and 33.68% in the warm season feeding group and cold season feeding group, respectively. Actinomycetes accounted for 2.78% and 2.47% in the warm season feeding group and the cold season feeding group, respectively. Patellar bacteria accounted for 3.81% and 1.00% in the warm season feeding group and cold season feeding group, respectively. None of the remaining phyla accounted for more than 1%. The cyanobacteria portal warm-season supplementation group was significantly higher than that of the cold season supplementation group (p< 0.01). The Bacteroides portal cold season feeding group was significantly higher than that of the warm season feeding group (p< 0.05), the patella bacterial portal warm season feeding group was significantly higher than the cold season feeding group (p< 0.05), and the spirochete portal cold season feeding group was significantly higher than that of the warm season feeding group (p< 0.05).
This is shown in Table 7, A total of 194 genera were detected at the genus level, and their dominant genera were Christensen _R-7, unclassified rumen bacteria, Prevobacterium species, Rikenia species, uncultured bacteria, NK4A214 genus, rumen species, indeterminate species, uncultured and Webron's species. Among them, except for the cold season supplementary feeding group, the proportion of Wehronia species did not exceed 1%, and the proportion of other dominant bacteria in the warm season supplementary feeding group and the cold season supplementary feeding group exceeded 1%. The cold-season supplementation group of Prevobacterium and the uncultured species (uncultured_bacterium) were significantly higher than those in the warm-season supplementation group (p< 0.05). The warm season feeding group of Rumen and Wehronia was significantly higher than that of the coldseason supplementation group (p< 0.05).
We performed LEFSE analysis on rumen fluid from two groups to detect changes in the composition of bacterial taxa. The LDA value is greater than 2, and the significant difference between the two groups is shown in Figure 2. In addition, when the microbial communities of grazing yaks with different seasons were compared with the same concentrate level, the most abundant bacterial genera in the warm season supplemental feeding group (WGYS) wasVeillonellacace_UCG_001, Candidatus_Saccharimonas, and UCG_001. The most abundant bacterial phylum were Firmicutes and Patescibacteria. The most abundant genera of bacteria in the cold season supplementation group (GYS) were Prevotella and Prevotellaceae_UCG_003, and the most abundant bacterial phylum was Bacteroidota.
This is shown in Figure 3, The yak rumen samples from this trial had 32 gene families at KEGG2 levels. Five of the two groups had significant differences (p< 0.01). There were 7 significant groups in the two groups (p< 0.05). None of the remaining 19 groups were significant. In this experiment, the warm-season supplementation group of Celular community-prokaryores was significantly lower than that of the cold-season supplementation group. The transcription warm-season feeding group was significantly lower than the cold-season supplementation group. The biodegradation and metabolism of exogenous substances was significantly higher in the warm season feeding group than in the cold season feeding group. Nucleotide metabolism was significantly higher in the cold season feeding group than in the warm season feeding group. The cold season feeding group was significantly higher than the warm season feeding group. The Metabolism of cofactors and vitamins were significantly higher in the cold season group than in the warm season feeding group. The cold season feeding group of the immune system was significantly higher than that of the warm season feeding group (p< 0.05). The warm-season supplementation group of membrane transport was significantly higher than that of the cold-season supplementation group (p< 0.01). Metabolism of other amino acids was significantly higher in the warm season feeding group than in the cold season feeding group(p< 0.01). Biosynthesis of other secondary metabolites was significantly higher in the cold season feeding group than in the warm season feeding group(p< 0.01). Carbohydrate metabolism was significantly higher in the cold season feeding group than in the warm season feeding group(p< 0.01). The cold season feeding group of the digestive system was significantly higher than that of the warm season feeding group(p < 0.01).
In this experiment, 2.5kg of concentrate was fed to grazing yaks in both warm season and cold season, and the total weight gain and average daily weight gain of the warm season feeding group were significantly higher than those in the cold season feeding group (p< 0.05), and the total weight gain of the warm season supplementary feeding group increased by 82.39% and the average daily weight gain increased by 82.61% compared with the cold season supplementary feeding group. The warm season feeding group gained 0.38kg/head per day compared with the cold season supplementary feeding group. This is similar to the results of Johnson [11] and Holley et al [12]. It showed that the concentrate of yak supplementation in Qinghai-Tibet region in the cold season was higher than 2.5kg/head per day to reach the growth state of the warm season, and the average net profit per head of the warm season supplementary feeding group increased by 214.84% compared with the cold season supplementary feeding group. That is, the net profit increased by 700.29 yuan per head. However, in the cold season, each yak was fed 2.5kg of concentrate every day, and the net profit within two months of the cold season was also 325.95 yuan per head. Therefore, supplementing 2.5kg of concentrate in the cold season can not only increase the breeding efficiency, but also make great sense for yaks to pass the adaptation period in the second year
The main sources of ruminants are volatile fatty acids such as acetic acid and propionic acid [9], which in turn are produced by the fermentation of glycogenic substances in feed by rumen fluid. Due to seasonal changes, the decrease in fiber content of cold season forage leads to a decrease in pH of grazing yaks in the cold season. This is similar to the results of the study of Pang Kaiyue et al [13]. Rumen ammonia nitrogen is a substance that synthesizes microbial proteins. It is synthesized by rumen microorganisms using nitrogen in feed. Its concentration is related to the rate at which rumen microorganisms break down and utilize nitrogen. The other studies have shown that the higher the protein content in the diet, the lower the concentration of ammonia nitrogen. In this experiment, the ammonia nitrogen cold season feeding group was significantly higher than that in the warm season feeding group (p< 0.01). The microbial protein warm season feeding group was higher than the cold season feeding group, and the difference was not significant (p>0.05). This indicated that the dietary protein content of the warm season feeding group was higher than that in the cold season feeding group, and the microbial protein synthesis was accelerated. This is consistent with the results of others. The percentage of acetic acid increases with the percentage in starch and the percentage of propionic acid changes with the percentage of fiber. In this experiment, the cold season supplementation group of volatile fatty acids acetic acid was significantly higher than that of the warm season feeding group (p< 0.05), and the propionic acid warm season feeding group was significantly higher than that of the cold season supplementation group (p< 0.01). This indicates that the content of starch substances in the cold season feeding group is lower than that in the warm season feeding group. The fiber feeding group was lower in the warm season feeding group than the cold season feeding group. The level of concentrate supplementation in both groups was the same, which proved that the nutritional value of coarse material in the cold season was lower than that of coarse material in the warm season. Wang Xianjing et al [14] showed that increasing the percentage of concentrate with a subsequent increase in propionic acid concentration improved animal performance. This is consistent with the results of this trial. This shows that warm season supplementation is better than cold season supplementation for feed utilization. Isobutyric acid is closely related to the growth of rumen microorganisms, and the increase in isobutyric acid content is conducive to the growth of rumen microorganisms and thus affects rumen fermentation. In this experiment, the warm season feeding group of isobutyric acid was significantly higher than that of the cold season feeding group. Valeric acid butyrate was related to fat synthesis, and the valeric acid in the warm season feeding group was significantly higher than that in the cold season feeding group. The butyric acid content was higher in the warm season feeding group than the cold season feeding group.It shows that warm season supplementation is more conducive to rumen fermentation than cold season supplementation. However, studies have shown that cold season supplementation also promotes rumen fermentation. Rumen fermentation is related to the level of concentrate, so it is beneficial to rumen fermentation that can be moderately increased without affecting the health of yaks.
In this experiment, the effect of the same supplementary feeding level on the serum biochemical indexes of grazing yaks in different seasons, Serum cholesterol and total triglycerides are indicators of fat metabolism [15-16]. The warm-season cholesterol feeding group was higher than that of the cold-season supplementation group, and the total triglyceride cold-season supplementation group was significantly higher than that of the warm-season supplementation group (p< 0.05), This is not related to Kaplan, B et al the results were similar [17]. Abnormal liver function is primarily related to serum alanine aminotransferase and glutamin aminotransferase[18]. In this trial, the glutamin aminotransferase warm-season supplementation group was significantly higher than that in the cold-season supplementation group (p< 0.01), indicating that concentrate supplementation should be appropriately increased in the cold season. The amount of total protein and albumin in serum reflects the level of antibodies and is also related to protein utilization and metabolism. The total protein warm season feeding group was significantly higher than that of the cold season feeding group (p< 0.05), and the albumin warm season feeding group was significantly higher than that of the cold season feeding group (p< 0.05). It was shown that compared with cold season supplementation, the level of antibodies and protein utilization rate were improved. Urea nitrogen is associated with protein content and renal protein metabolism and is an indicator of renal disease surveillance. Therefore, it is recommended to moderately increase the amount of concentrate supplementation for grazing yaks in the cold season, which is conducive to the health of yaks.
16S rDNA high-throughput sequencing technology has a better understanding of the effect of yak feeding with the same concentrate level on the rumen microbiota in different seasons. Microbial diversity is an indicator for measuring the quality of rumen microorganisms, mainly including six indicators. Their size is an important parameter to measure the richness and diversity of the flora. This study showed that the number of unique OTUS was 12 more OTUs in the warm season feeding group than in the cold season feeding group. Domestic scholars found [19] that with the increase of the proportion of concentrate, the Chao1 index, Shannon index and genealogical diversity index decreased significantly, while the difference in Simpson index was not significant. Studies such as Pang Kaiyue [20] have shown that increasing the ratio of grain to thickness reduces the diversity and richness of rumen bacteria. In the results of this trial, there was no significant difference in the six indicators, which may be due to the fact that the level of concentrate supplementation in the warm season group and the cold season feeding group were the same.
Through the study of ruminant rumen microorganisms, scholars have generally reached a consensus that Firmicutes and Bacteroides are the dominant phylums of ruminant rumen microorganisms [21-22]. It is mainly involved in the decomposition of crude fiber and other fibrous substances in ruminant diets is the phylum Firmicutes; Phylum Bacteroides is mainly involved in the degradation of non-fibrous substances such as starch[23-24]. These two phylums play a particularly important role in energy metabolism in ruminants. At the phylum level, a total of 21 phylums were detected, Firmicutes and Bacteroides were the dominant phylums, and Firmicutes accounted for 64.61% and 60.69% in the warm season and cold season feeding groups, respectively. Bacteroides accounted for 25.93% and 33.68% in the warm season feeding group and cold season feeding group, respectively. Actinomycetes accounted for 2.78% and 2.47% in the warm season feeding group and the cold season feeding group, respectively. The proportion of actinomycetes in the rumen was second only to Bacteroides in the cold season supplementation group. Patellar bacteria accounted for 3.81% and 1.00% in the warm season feeding group and cold season feeding group, respectively. The proportion of patellar bacteria in the phylum is higher than 1%. None of the remaining phylums accounted for more than 1%. The bacteria contained in the phylum Cyanobacteria have the function of producing oxygen[25], and the warm season feeding group of cyanobacteria is significantly higher than that of the cold season supplementation group (p< 0.01), indicating that cold season supplementation is more conducive to the creation of rumen anaerobic environment and thus rumen fermentation than warm season supplementation. The Bacteroides portal cold season feeding group was significantly higher than that in the warm season feeding group (p< 0.05), which was consistent with the results of Liu Xiu [11] and others, indicating that the proportion of starch feed in the cold season feeding group was higher than that in the warm season feeding group. Due to the consistency of the supplemental concentrate, it may be that the feed intake of yaks during grazing in the cold season is reduced compared with that during supplementation in the warm season. Therefore, grazing in the cold season is to ensure the production efficiency of yaks, and the amount of supplementary feeding should be appropriately increased in the cold season. The patellar bacterial phylum warm-season supplementation group was significantly higher than that of the cold-season supplementation group (p< 0.05), and the results of Zhou Wenbin [26] showed that the patellar bacterial phylum was lower than that in the liver cancer group, indicating that patellar bacterial phylum may be related to pathogenic factors, indicating that cold-season supplementation may be more conducive to the health of yaks. Li Lingxuan [27] and other studies have shown that increasing the nutritional level of the diet increases the relative abundance of the spirochete phylum. In this trial, the spirochete portal cold season feeding group was significantly higher than the warm season feeding group (p< 0.05). Studies have shown that the spirochete phylum is related to the energy metabolism of ruminants , indicating that the energy metabolism in the cold season is more vigorous than that in the warm season, so to maintain production efficiency in the cold season, the amount of concentrate supplementation should be appropriately increased.
A total of 194 genera were detected at the genus level, and their dominant genera were Christensen _R-7, unclassified rumen bacteria, Prevobacterium species, Rikenia species, uncultured bacteria, NK4A214 genus, rumen species, indeterminate species, uncultured and Webron's species. Among them, except for the cold season supplementary feeding group, the proportion of Wehronia species did not exceed 1%, and the proportion of other dominant bacteria in the warm season supplementary feeding group and the cold season supplementary feeding group exceeded 1%. Gao Yufei [28] The study found that increasing the dietary energy level increased the relative abundance of Prévoria spp., and the cold season feeding group of Prévoria in this experiment was significantly higher than that in the warm season feeding group (p< 0.05). The proportion of concentrate in the warm season supplementary feeding group was higher. The cold season feeding group of uncultured mycetes (uncultured_bacterium) was significantly higher than that in the warm season feeding group (p< 0.05). Domestic scholars found [29-30] that the abundance of rumen in ruminant rumen was closely related to feed fibers, and in this experiment, the rumen was significantly higher in the warm season feeding group than the cold season feeding group (p< 0.05), indicating that the warm season feeding group had more forage. The warm-season supplementation group of Willon's species was significantly higher than that of the cold-season supplementation group (p< 0.05), and the concentration of Willon's species is one of the factors that promote the growth of cancer cells, and its elevated concentration is not conducive to the health of animals. There were no significant differences in the other groups. In summary, supplementation in the cold season is to appropriately increase the proportion of concentrate supplementation, and appropriately reduce the proportion of concentrate supplementation in the warm season, which is conducive to the growth and health of yaks in the cold and warm season.
PICRUSt was used to predict the metabolic pathway and function of rumen microorganisms, and there were 32 gene families in yak rumen samples at KEGG2 level. Five of the two groups had significant differences (p< 0.01). There were 7 significant groups in the two groups (p< 0.05). None of the remaining 19 groups were significant. Carbohydrate metabolism The most abundant metabolic pathway in this test, followed by Metabolism of cofactors and vitamins, amino acid metabolism and metabolism of terpenoids and polyketides, accounted for more than 10% in the warm season feeding group and cold season supplementary feeding group. The main metabolic pathways are closely related to the metabolism of sugars, amino acids and fats. These are essential for the survival, growth, and reproduction of animal gastrointestinal microbial communities [31]. Energy is an important part of feed, the most critical factor in animal feed intake, and an important substance for animals to survive, carry out life activities and reproduce. In this experiment, the warm-season supplementation group of Celular community-prokaryores was significantly lower than that of the cold-season supplementation group. The transcription warm-season feeding group was significantly lower than the cold-season supplementation group. The biodegradation and metabolism of exogenous substances was significantly higher in the warm season feeding group than in the cold season feeding group. Nucleotide metabolism was significantly higher in the cold season feeding group than in the warm season feeding group. The cold season feeding group was significantly higher than the warm season feeding group. The Metabolism of cofactors and vitamins were significantly higher in the cold season group than in the warm season feeding group. The cold season feeding group of the immune system was significantly higher than that of the warm season feeding group (p< 0.05). The warm-season supplementation group of membrane transport was significantly higher than that of the cold-season supplementation group (p< 0.01). Metabolism of other amino acids was significantly higher in the warm season feeding group than in the cold season feeding group. Biosynthesis of other secondary metabolites was significantly higher in the cold season feeding group than in the warm season feeding group. Carbohydrate metabolism was significantly higher in the cold season feeding group than in the warm season feeding group. The cold season feeding group of the digestive system was significantly higher than that of the warm season feeding group. The above shows that the same concentrate level of supplementation in the cold and warm season is vigorous in the cold season, and the metabolic intensity in the warm season is generally not as good as the metabolic intensity in the cold season. This showed that the demand for various nutrients in the cold season feeding group was higher than that in the warm season feeding group. Therefore, the amount of each nutrient should be appropriately increased during supplementary feeding in the cold season
This experiment supplemented each yak grazing in the cold and warm seasons with 2.5kg/d of concentrate. The experiment showed that the average daily gain of yaks from warm season to cold season also significantly decreased, inhibited rumen fermentation, decreased yak antibody levels, and abundance of rumen microorganisms, but increased the concentration of pathogenic factors. The abundance of beneficial microorganisms was reduced at the phylum and genus levels. At the KEGG level, it was shown that the cold season supplement group had strong metabolism and a higher demand for nutrients than the warm season supplement group.
The data and materials are authentic and reliable, without any conflict of interest
We thank Dr. Dongwen Dai of Ningxia University for doing the experimental design and Master Kaiyue Pang of Qinghai University for data analysis. We thank Researcher Chai Sha Camel, Dr. Shuxiang Wang and Dr. Yingkui Yang from the College of Animal Husbandry and Veterinary Medicine, Qinghai University for their financial support of this experiment.
All experiments in this study were performed following the approved guidelines of the Regulation of the Standing Committee of Qinghai People’s Congress. All experimental protocols and the collection of samples were approved by the Ethics Committee of Qinghai University under permission no. SL-2021027. This study complied with ARRIVE guidelines.
All data belongs to me and does not require authorization from third parties.

![]()

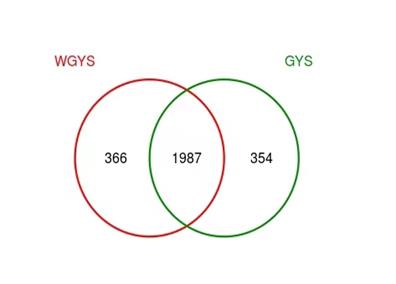 |
| Figure 1: Venn diagram |
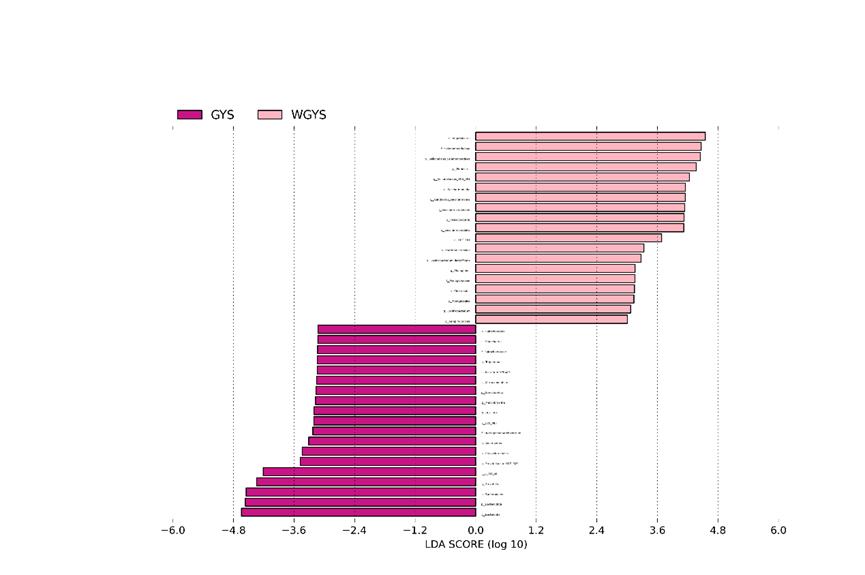 |
| Figure 2: LEFES analysis |
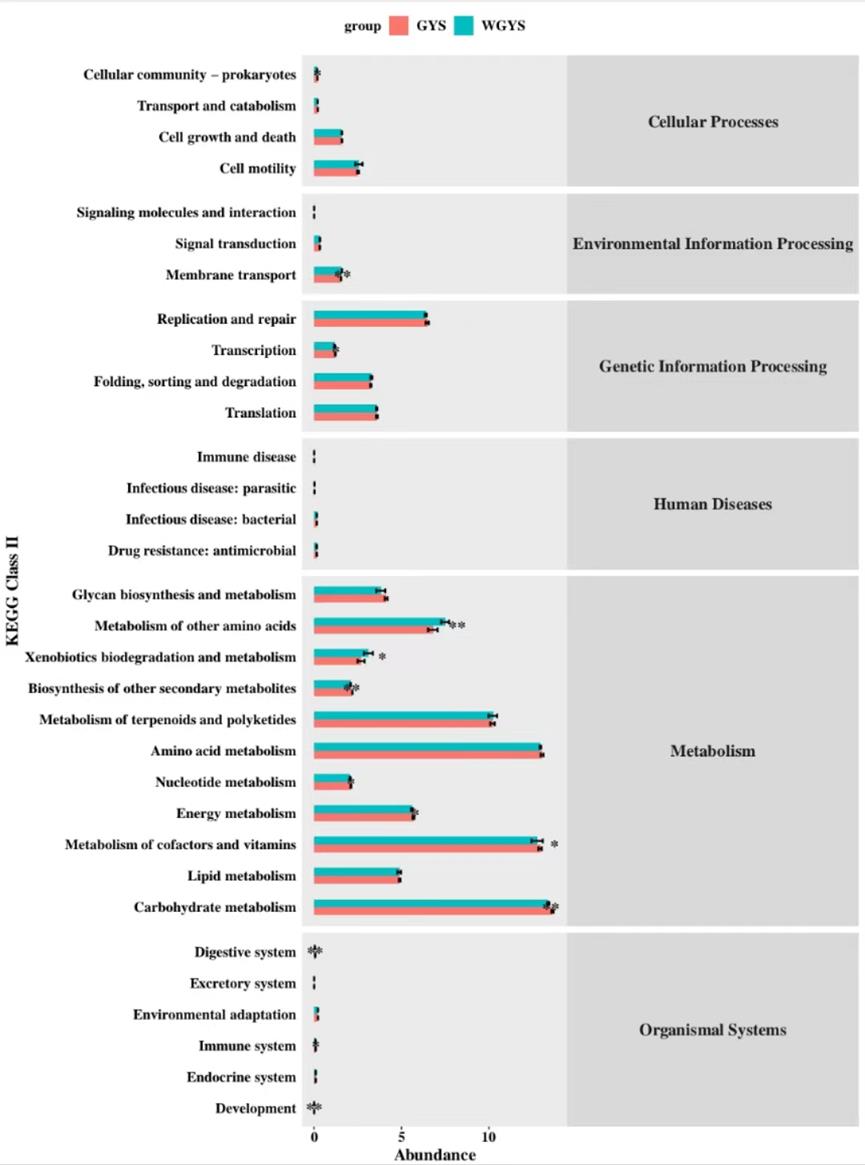 |
| Figure 3: KEGG |
Items |
Pasture |
Pasture |
Concentrate supplement(%) |
Ingredients |
|
|
|
Corn |
|
|
44.27 |
Wheat bran |
|
|
12.06 |
Wheat |
|
|
12.88 |
Soybean meal |
|
|
4.98 |
Cottonseed meal |
|
|
12.28 |
Ca(HCO3)2 |
|
|
8.57 |
NaHCO3 |
|
|
1.22 |
NaCl |
|
|
0.87 |
Premix1) |
|
|
0.87 |
Total |
|
|
100.00 |
Nutrient levels2) |
4.59 |
8.66 |
12.67 |
CP(%) |
4.67 |
11.35 |
18.76 |
EE(%) |
3.12 |
2.53 |
2.63 |
NDF(%) |
57.26 |
54.16 |
16.03 |
ADF(%) |
38.64 |
26.74 |
7.83 |
Ca(%) |
0.77 |
2.15 |
0.60 |
P(%) |
0.03 |
0.08 |
0.73 |
Items |
WGYS |
GYS |
SEM |
p-value |
Total weight gain (kg) |
50.04a |
27.45b |
2.707 |
p<0.001 |
Average daily gain in weight (kg/d) |
0.84a |
0.46b |
0.045 |
p<0.001 |
Material-to-weight ratio |
2.98 |
5.43 |
|
|
Average net profit (RMB: Chinese Yuan) |
1026.24 |
325.95 |
|
|
Items |
WGYS |
GYS |
SEM |
p-value |
pH |
6.88 |
6.86 |
0.080 |
0.862 |
NH3-N |
10.20B |
13.58A |
0.550 |
P<0.001 |
MCP |
2.49 |
2.32 |
0.196 |
0.357 |
Acetic acid |
42.90b |
45.99a |
1.470 |
0.045 |
Propionic acid |
19.16A |
13.33B |
0.680 |
P<0.001 |
Isobutyric acid |
1.03A |
0.65B |
0.060 |
P<0.001 |
Butyric acid |
8.94 |
7.78 |
0.646 |
0.090 |
Isovaleric acid |
2.12A |
0.89B |
0.069 |
P<0.001 |
Valeric acid |
2.04A |
1.12B |
0.046 |
P<0.001 |
Acetic acid/propionic acid |
2.27B |
3.45A |
0.102 |
P<0.001 |
Items |
WGYS |
GYS |
SEM |
p-value |
Glucose |
4.67 |
4.76 |
0.190 |
0.672 |
Cholesterol |
2.30 |
2.19 |
0.106 |
0.286 |
Total protein |
78.51a |
70.93b |
2.125 |
0.017 |
Albumin |
36.53a |
33.93b |
1.199 |
0.042 |
Globulin |
41.98 |
36.99 |
2.713 |
0.079 |
Urea nitrogen |
5.17B |
6.14A |
0.263 |
0.001 |
Alanine aminotransferase |
44.83A |
37.08B |
2.237 |
0.002 |
Glutamate aminotransferase |
93.17 |
87.00 |
6.197 |
0.331 |
Alkaline phosphatase |
208.25 |
208.67 |
8.955 |
0.963 |
Items |
WGYS |
GYS |
SEM |
P |
Chao1 index |
1782.28 |
1809.07 |
59.937 |
0.671 |
goods_coverage index |
0.99 |
0.99 |
0.0 |
|
observed_species index |
1506.98 |
1486.95 |
58.063 |
0.345 |
PD_whole_tree |
118.86 |
116.74 |
3.824 |
0.599 |
Shannon index |
8.36 |
8.32 |
0.202 |
0.822 |
Simpson index |
0.99 |
0.99 |
0.0 |
|
Items |
WGYS |
GYS |
SEM |
p-value |
Firmicutes phylum |
64.61 |
60.69 |
1.043 |
0.227 |
Phylum Bacteroides |
25.93B |
33.68A |
1.774 |
0.005 |
Actinomycete phylum |
2.78 |
2.47 |
0.626 |
0.638 |
Phylum of patellar bacteria |
3.81a |
1.00b |
1.027 |
0.034 |
Phylum of verrucous microbacteria |
0.94 |
0.95 |
0.359 |
0.979 |
Phylum Desulfobacillus |
0.47 |
0.51 |
0.120 |
0.765 |
Phylum Spirochaetota |
0.21b |
0.51a |
0.093 |
0.016 |
Phylum Cyanobacteria |
0.29A |
0.11B |
0.046 |
0.009 |
Proteobacteria phylum |
0.19 |
0.15 |
0.078 |
0.654 |
Phylum Chlorophylum |
0.22 |
0.103 |
0.080 |
0.116 |
Items |
WGYS |
GYS |
SEM |
p-value |
Christensenellaceae_R-7_group |
12.69 |
14.58 |
3.143 |
0.519 |
uncultured_rumen_bacterium |
8.84 |
10.73 |
0.992 |
0.106 |
Genus Prevo |
5.55b |
9.58a |
1.458 |
0.026 |
Genus Phyllus |
7.27 |
6.70 |
0.838 |
0.518 |
uncultured_bacterium |
6.12b |
7.02a |
1.327 |
0.030 |
NK4A214_group |
5.24 |
6.45 |
0.810 |
0.185 |
Rumen species |
5.43a |
3.62b |
1.828 |
0.018 |
unidentified |
4.72 |
3.90 |
0.993 |
0.440 |
uncultured |
3.79 |
2.94 |
0.652 |
0.243 |
Genus Wersonia |
4.20a |
0.95b |
0.974 |
0.016 |




































