Open Access
Research Article
Max Screen
ISSN: 2348-9790
Copyright: © 2023 Jia Yin1. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
The purpose of this study was to clarify the response of Ningxiang pig against pathogenic enterotoxigenic Escherichia coli (ETEC) during the weaning period. Sixteen weaned Ningxiang pigs, were selected from one litter at 28 days and randomly allocated into two groups, with or without oral administration of ETEC (109 colony forming units/piglet/d) for two consecutive days. Blood and intestinal samples were collected after sacrifice to investigate the influence for systemic and gastrointestinal tract influences. We profiled growth performance, blood parameters, jejunal immunity gene expression, microbiota composition and intestinal metabolites in weaning Ningxiang piglets using the ETEC challenge model. The piglets received ETEC had downregulated mean corpuscular volume (MCV) and mean corpuscular hemoglobin (MCH) levels, compared to the controls. The transcriptional levels of gclc, gclm and tlr2 genes related to antioxidant and immunity status in the jejunum were highly expressed after ETEC challenge. Meanwhile, 16S rRNA sequencing in colon showed a significantly high abundance of Oxalobacter in the ETEC group. The concentrations of short chain fatty acids (SCFAs) and total bile acids were suppressed after ETEC challenge. In swine industry, pathogenic ETEC commonly leads to enteric inflammation and diarrhea. During the long-term domestication, Ningxiang pigs, with their unique hereditary properties, are a well-known indigenous breed in China. The imbalance of redox and their immune systems might interact with the metabolic function of gut microbiota after ETEC challenge. Overall, we described the impact of ETEC on Ningxiang pigs, shedding light on the protection of indigenous swine breeds.
Keywords: ETEC; weaning stress; Ningxiang pig; gut microbiota
Post-weaning diarrhea in piglets is associated with enteric pathogen infections [28]. Pathogenic ETEC is related to the worldwide spread of antibiotic-resistant genes among various ecological niches [60]. The ETEC W25K strain produces the F4 fimbriae, lipopolysaccharide, heat-stable enterotoxin, and heat-labile enterotoxin [42,62]. In diarrhea piglets, proteome analysis indicated that ETEC W25K suppressed jejunal immune responses. In porcine intestinal epithelial cells, ETEC infection could induce proinflammatory responses via mitogen-activated protein kinases and the nuclear factor kappa B pathway [48,67,42,43].
Though less productive than commercial breeds, indigenous swine breeds possess valuable genetic and ecological qualities [17].The Yorkshire × Landrace pig is sensitive to porcine circovirus type 2 with severe clinical features, and yet the indigenous Laiwu pig shows immunological stress resistance to the virus infection, partly depending on the up-regulated SERPNA1 gene [50,29].Congjiang pig takes advantage of microbiota-derived gassericin A to prevent post-weaning diarrhea [19]. Ningxiang pigs, which have high intramuscular fat meat [9], have been a well-known native livestock resource and geographic representative of agricultural products for over a thousand years [15]. In addition to the pleasant meat flavor, Ningxiang pigs can also serve as a meaningful breed resource for disease resistance [64,17].c
Although researchers have been aware of the consequences of ETEC infection in crossbred commercial pig [4], the ETEC-induced alteration in native pigs has not been clarified to date. We investigated whether Ningxiang pigs are resistant to ETEC challenge by analysis of the growth performance, fecal score, blood indexes, gut morphology, intestinal microbiota and metabolites. The findings revealed that ETEC led to alterations of oxidation balance and immune compromise, which were accompanied by decreased concentrations of total bile acids in ileum and SCFA in colon. This investigation offered insights into further study pathogenicity of ETEC and host response in Ningxiang pigs.
The ETEC W25K strain (O149:K91, K88ac; LT, ST, EAST) [42], was preserved in 25% (v/v) glycerol broth at refrigerator. The ETEC strain was streaked and cultured aerobically at 37 °C overnight. Single colonies from the agar plate were picked and cultured in Luria-Bertani (LB) liquid medium at 37 °C overnight. The bacterial concentration was counted by the colony forming unit(CFU) after serial dilution. Next, sterile LB liquid was served as a diluent for achieving the 1 × 108 CFU/mL concentration. The solution was transported with ice packs to the farm site.
The piglet trial procedures were approved by the Animal Care and Use Committee of Hunan Normal University (Permit Number: 2020023). Sixteen Ningxiang piglets, including male and females, were selected from one litter at the Chuweixiang farm in Changsha city. They were weaned at 28 days with an average initial body weight (5.49 ± 0.33 kg). All piglets were randomly allotted to two treatments and housed individually. The control group: The piglets were given 10 mL of sterile LB liquid broth by gavage at 31 and 32 days. The ETEC group: The piglets were given the 10 mL ETEC broth at a dose of 1 × 108 CFU/mL by gavage for two consecutive days. A corn–soybean meal basal piglet diet was formulated to meet the Nation Research Council (NRC, 2012) recommendations. Piglets had free access to water and were fed ad libitum.
Fecal consistency (0 = normal feces, 1 = soft feces, 2 = mild diarrhea, and 3 = watery and projectile diarrhea) was scored as described previously [66], at 31, 32, 33 and 34 days.
All piglets were fasted overnight, euthanized through intravenous injection with 150 mg/kg body weight pentobarbital (Kela, Hoogstraten, Belgium), and then were sacrificed at 35 days. Approximately 10 mL of blood was collected from each piglet by venipuncture at the anterior vena cava, followed by centrifugation at 3,000 g at 4 0C for 15 min.
Approximately 2 cm of intestinal tissues was cut from the ileum, duodenum, jejunum and colon. Paraformaldehyde solution was used to fix tissues for morphology analysis. The mucosae in jejunal segment were separated for real-time PCR. Intestinal digesta from the ileum and colon were immediately transported in liquid nitrogen for the 16S rRNA gene amplicon sequencing and volatile fatty acid analysis.
Whole blood was collected into tubes containing ethylenediaminetetraacetic acid dipotassium salt dihydrate. The blood samples were mixed thoroughly, before determining the routine blood parameters using an automatic blood analyzer Advia 120. (Bayer HealthCare, Tarrytown, NY, USA). The concentrations of hemoglobin (HGB), hematopoietic cell transplantation (HCT), white blood cell (WBC), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), meanc orpuscular volume (MCV), mean platelet volume (MPV), plateletcrit (PCT), platelet distribution width (PDW), platelets (PLT) and red blood cell (RBC) were analyzed. The distribution width of red blood cells (RDW-CV) and average distribution width of red blood cells (RDW-SD) were also evaluated.
Besides, serum samples were analyzed using a TBA-120FR Automatic Biochemistry Radiometer (Hitachi Co., Tokyo, Japan) at the hospital of Hunan Normal University. According to the instructions, serum parameters were determined using commercial kits (Jiancheng Bioengineering Institute, Nanjing, China). The considered parameters were α-amylase (α-AMY), albumin (ALB), alanine aminotransferase (ALT), aspartate aminotransferase (AST), the postoperative ratio of albumin to blood glucose (A/G), blood urea nitrogen (BUN), alkaline phosphatase (ALP), cholesterol (CHO), creatinine (CREA), direct bilirubin (DBIL), globulin (GLO), glucose (GLU), indirect bilirubin (IBIL), immunoglobulin A (IgA), lactate dehydrogenase (LDH), low density lipoprotein cholesterol (LDL-C), high density lipoprotein cholesterol (HDL-C), total bilirubin (TBIL), triglyceride (TG) and total protein (TP).
Each sample was cut longitudinally at the middle of intestinal segments. Villus height and crypt depth were determined based on a previous study [20]. Briefly, fixed intestinal samples were embedded in paraffin. Sections of 5 μm were cut and stained with hematoxylin-eosin. Imagines were captured under a light microscope Leica DM3000 (Leica Microsystem, Wetzlar, Germany). Morphological measurements were measured in Image Pro-Plus Software version 6.0 (Media Cybernetics, Maryland, USA) without knowledge of treatment.
The SCFAs concentrations in colon were quantified, as a previous protocol [65]. Briefly, each sample was weighted and thoroughly diluted with precooled distilled water, followed by centrifugation at 10,000 g for 15 min. The supernatant was collected and mixed with 25% metaphosphoric acid at a ratio of 1:4 (m/v) at 4 °C overnight. After precipitate removal using a filtration membrane, the one microliter of supernatant sample was injected into a gas chromatograph system with DB-FFAP column (Agilent Technologies, Delaware, USA). The injector and detector temperatures were set as 250 °C and 280 °C, respectively.
The intestinal digesta were weighed and homogenized in distilled water. Using a commercial assay kit (Jiancheng, Nanjing), the levels of total bile acids in the ileum and colon were measured at a wavelength of 405 nm and normalized to the weight of intestinal digesta.
Total RNA was extracted by the TRIZOL reagent (Qiagen, CA, USA) and DNase. The quantity and quality of RNA were checked through electrophoresis and ultraviolet spectroscopy (Thermo Fisher Scientific, Delaware, USA), followed by reverse transcription using iScriptTM cDNA Synthesis Kit (Bio-Rad, Hercules, CA). The ABI 7900HT Fast Real-Time PCR System (Carlsbad, CA, USA) was used for real-time quantitative PCR analyses at a final volume of 20 μL. All PCR reactions were run in triplicate with the following conditions: 95 °C for 30 secs, followed by 35 cycles of 95 °C for 3 sec and 58 °C for 30 sec. The β-actin was used as an internal control. The relative expressions of the interested genes were quantified by the 2(-Delta Delta C(T)) method [31]. All reactions were run in triplicate for each sample. The primer sequences were as follows, gclm forward: 5’- GATGCCGCCCGATTTAACTG-3’, gclm reverse: 5’-ACAATGACCGAGTACCGCAG-3’; gclc forward: 5’- CAAACCATCCTACCCTTTGG-3’, gclc reverse: 5’-ATTGTGCAGAGAGCCTGGTT-3’; tlr2 forward: 5’-GTTCACGCATTTCCGCAGTT-3’, tlr2 reverse: 5’- CTTTGTGGACAGCATGGGTCTT-3’; actin forward: 5’- AGTTGAAGGTGGTCTCGTGG-3’, actin reverse: 5’-TGCGGGACATCAAGGAGAAG-3’.
Microbial DNA from colonic digesta was extracted using the intestinal contents DNA extraction kit (TIANNampTIANGEN, China). A pair of primers were used to amplify the 16S rRNA hypervariable region V3-V4 as 338F (5'- ACTCCTACGGGAGGCAGCAG-3') and 806R (5'-GGACTACHVGGGTWTCTAAT-3'). The polymerase chain reaction mixtures in a volume containing TransStart FastPfu buffer and DNA polymerase.
Amplicons were sequenced by an Illumina MiSeq PE300 platform (Illumina, San Diego, USA) with primers and barcodes, followed by an adjustment of the sequence direction [12]. Using the UPARSE software, operational taxonomic units (OTUs) were calculated at 97% sequence similarity [11]. The taxonomy of the representative sequence was against the SILVA version 132 database [39]. The alpha diversity and the bata diversity were calculated using QIIME2 [6].
Data in the study were processed and visualized by SPSS software version 19.0 (IBM Corp., Chicago, USA) and GraphPad Prism software version 8.3 (San Diego, CA, USA). The results were presented as means ± SEMs. For the statistical analyses, probability values less than 0.05 after unpaired Students two-tailed t-test were considered significant. The differences were defined as trends at probability values less than 0.1. n.s., indicates not significant.
Sixteen Ningxiang piglets were randomly separated into a control group and an ETEC group (Figure 1A). In comparison to the control group, the final body weight in the ETEC treated group showed no significant difference (P = 0.15) (Figure 1B-C). There was also no significant difference in the diarrhea index or diarrhea rate between the two groups (Figure 1D-E).
The morphologies of different intestinal segments were measured by hematoxylin-eosin staining for the duodenum, jejunum, ileum and colon to examine the impact of ETEC challenge on intestinal development (Figure 2-3). Ileal crypt depth showed a decreased trend in the ETEC group (P = 0.062) (Figure 2I and Figure3C).
Haematological analysis was detected by blood routine and biochemistry parameters. ETEC challenge significantly suppressed the concentrations of MCV in the blood (P < 0.05) (Figure 4A). It is also worthwhile that there was a decreased trend in the ETEC group for MCH (P = 0.053), compared with the control group (Figure 4B). Nevertheless, there was no significant difference for other parameters in blood routines between the two groups (Table S1).
Besides, serum concentrations of α-AMY, ALB, ALP, ALT, A/G, BUN, CHO, CREA, DBIL, GLO, GLU, HDL-C, IBIL, IgA, LDH, LDL-C, TBIL, TG and TP between the control group and ETEC group were comparable (Table S2). There was an increasing trend for serum AST in the ETEC group (P = 0.097) (Figure 4C).
ETEC infection was previously reported to induce intestinal inflammation and disturb the prooxidant-antioxidant balance in swine [53]. Next, some gene expressions related to inflammation and antioxidant ability were analyzed by real-time PCR (Figure 4D-F). It revealed that ETEC supplementation positively regulated the gene expressions of gclm (P = 0.076), gclc (P = 0.098) and tlr2 (P < 0.05) in the jejunum mucosa of Ningxiang piglets.
Changes in the colonic microbiota after ETEC stimulation were monitored by 16S rRNA gene sequencing. In the alpha diversity indexes, Chao1, Goods coverage index, observed features, Pielou evenness index, Shannon-Wiener diversity index and Simpson diversity index had slight variations (Figure 5A). At the genus level, PCoA demonstrated that gut bacteria showed no differences between ETEC group and the control group by Jaccard and Anosim pairwise test (P = 0.236) (Figure 5B). Furthermore, the taxonomy analysis revealed that the relative abundance of Oxalobacter genus markedly increased in the ETEC group (P < 0.05),compared to the control group (Figure 5C).
To further investigate the effects on the colonic metabolites, we detected the SCFA concentrations in piglets. Among volatile fatty acids, the ETEC challenge decreased the levels of acetic acid (P < 0.05), propionic acid (P < 0.05) and butyric acid (P = 0.085) in the colon digesta (Figure 6A-C). However, for isobutyric acid, isovaleric acid and valeric acid, the ETEC challenge did not cause any statistical differences (Figure 6D-F).
Bile acids are essential physiological agents for regulating intestinal inflammation and host immune responses [10,16]. The result showed that ETEC challenge significantly reduced the concentration of total bile acids in the ileum (P < 0.05), but not in the colon (Figure 6G-H).
Escherichia coli is a universal pathogen contributing to diarrhea, immune responses and damaged tight junction function in swine [58,30]. We observed that ETEC administration did not significantly affect growth performance and diarrhea incidence in Ningxiang piglets, but also adversely affected immunology and antioxidant potential. To a certain extent, ETEC challenge inhibited the levels of MCV and MCH by the routine blood test, whereas, the concentration of serum AST displayed a slightly increased trend in the ETEC group.
Hematological cells are composed of leukocytes, erythrocytes and platelets [55]. During pathogen infection, associations between hematological cells and the immune response were widely reported [34, 32]. Notably, erythrocytes have highly antioxidant ability. Under oxidative stress, reactive oxygen species were removed [36]. MCV means the average size of erythrocytes. MCH is an estimate of the amount of hemoglobin in an average erythrocyte. In view of the reductions of MCV and MCH, ETEC might stimulate the local immune system and pose a threat to the redox balance for piglets. When sows were fed on selenomethionine, their antioxidant status was improved, which were characterized by the elevation of hematological MCV and MCH levels in piglets [13]. In humans, the immune system and hematopoietic system were impaired for workers exposed to benzene, accompanied with low MCV and MCH hematological levels [46].
AST is regarded as a serological indicator during liver injury, as the increasing AST level reflects the leakage from injured hepatocytes [37]. Addition of Clostridium butyricum-based probiotic agents in weaned piglets reduced the AST concentration in serum and improved growth performance [8].
The transcriptional analysis in Ningxiang pig displayed that the gene expression levels related to oxidative stress (gclc and gclm) and immunity (tlr2) were moderately increased in the jejunum with ETEC challenge. In the disease development of retinitis pigmentosa, the high relative expressions of gclc gene and gclm gene were widely observed [47,1]. Previous evidence indicated that piglets with ETEC infection were suffered from oxidative stress, which was characterized by excessive production of reactive oxygen species in the jejunum [27]. In agreement with our results, Roberts et al. reported that an opportunistic pathogen,Porphyromonas gingivalis, markedly increased the transcriptional levels of gclc and gclm in primary epithelial cells [45].
Typically for newborn piglets and weaning piglets, the development of intestinal immune system remains relative immature [7, 24]. TLR signal pathway participates in the activation of the innate immune system, which is initiated by the recognition of structural components of invading pathogens [33]. Intestinal epithelium cells express tlr2 to regulate inflammation [38]. In our study, high expressive level of tlr2 gene has been observed, which was consistent to a previous study that ETEC K88 challenge inhibited the negative regulators of TLR signaling [26]. For both human and mouse models, tlr2 is highly expressed in atherosclerotic lesions [5, 54]. Lactobacillus. plantarum was reported to ameliorate the ETEC induced inflammatory response in mice via TLR2 inflammatory signal pathway [21].
Intestinal microbiota are essential to maintain immune homeostasis of the host against infection [18]. Our study demonstrated that supplementation with ETEC did not significantly alter the diversity and the structure of colonic microflora, apart from the high abundance of Oxalobacter. Similarly, Xiao et al. revealed that, lipopolysaccharide had no effect on microbial diversity and richness, but partially changed the composition of gut microbiota in piglets, especially the upregulated abundance of Oxalobacter formigenes [63]. Oxalobacter formigenes is as a group of anaerobic bacteria that degrades oxalic acid [2]. However, the relationship between oxalic acid and ETEC still remains unclear.
The gut microorganisms decompose dietary nutrients and produce SCFAs and bile acids to modulate immune function [56,23,35]. Our results showed that the concentrations of acetate, propionate, and butyrate in colon were decreased in the ETEC group, which were accompanied with lower concentration of total bile acids in ileum, compared to the control group. The findings indicated that ETEC led to metabolic disorders and immune function impairment. During the weanling period, the piglets diets shift from high-fat milk to high-carbohydrate feed, resulting in more fermentable substrates and SCFAs reaching the colon [61]. SCFAs function as not only energy source to stimulate colonocytes growth, but also a signal in anti-inflammation to enhance the immunity [49,68]. For example, the abundance of Enterobacteriaceae is negatively correlated with the concentrations of undissociated acetic, propionic, and butyric acids in the caeca of broiler chickens [57]. Additionally, supplementation of formic and propionic acid was validated to alleviate the inflammatory response and reduce diarrhea incidence in weaning piglets challenged with ETEC [41]. Sun et al. found that dietary fibers could increase SCFAs and downregulate the TLR2 inflammatory signal pathway, which ameliorated lipopolysaccharide-induced intestinal damage in piglets [52].
Bile acids are abundant in the mammalian gut, where they effect host metabolism, innate immunity and adaptive immunity [61]. Ningxiang pig is a fat-type pig breed and is characterized by high back fat thickness [40]. Cholesterol in pork is a precursor to bile acid biosynthesis [14,51]. Bile acids have been considered to facilitate fat digestion. They are released into the intestine from the gallbladder, upon cholecystokinin stimulation [25,22]. Bile salts emulsify lipids and the majority of bile acids are reabsorbed at ileum. For instance, the pathogens of Clostridium perfringens and coccidia lead to necrotic enteritis in the poultry industry [59]. The development of chicken necrotic enteritis was characterized by reduced total bile acids in the ileal digesta [3]. In summary, the reduction of ileal total bile acids in our study of Ningxiang piglets might be related to immunocompromise and bile acid biosynthesis metabolism interfered by ETEC challenge.
Our study is the first attempt to investigate the host response of Ningxiang pig into pathogenic ETEC challenge at weaning period.Herein, ETEC led to oxidation imbalance and immunocompromise, which involved decreased MCV and MCH levels, and elevated gclc, gclm and tlr2 gene expression. Furthermore, the enhanced relative abundance of Oxalobacter genus was observed in colon of ETEC group, accompanied by the repressed concentrations of SCFAs in colon and total bile acids in ileum. Overall,though we have not provided clear mechanistic interactions between gut microbiota and the host immunological function yet, this study has expanded upon the understanding of pathogenic ETEC impact on native Ningxiang pigs, and shed light on the importance of protecting indigenous swine breeds in biomedical researches.
This work was supported by Prof. Wenkai Ren who donated the ETEC W25K strain. We are grateful for editorial assistance from the Xiaoni Cao, Yajie Wang and Wenjing Wang. We also thank Shanghai OE Biotec. Co., Ltd. and Majorbio Bio-Pharm Technology Co., Ltd. for the bioinformatics analysis.
This work was supported by the Guangdong Laboratory for Lingnan Modern Agriculture (NT2021005), Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project (TSBICIP-CXRC-038), the National Natural Science Foundation of China (32130099, 31700004), the Scientific Research Fund of Hunan Provincial Education Department (21A0060), Hunan Province College Students Research Learning and Innovative Experiment Project (S201910542041), Hunan Provincial Science and Technology Department (2019RS5001), Construction of Innovative Provinces in Hunan Province (2019RS3022) and the National Students Platform for Innovation and Entrepreneurship Training Program (201810542034, 2020056).
The raw reads were deposited into the NCBI database with accession number: PRJNA870752.
Competing of interests: Authors declare no conflict of interest.
Ethics approval: The experimental study using animals was approved by the Animal Care and Use Committee of the Hunan Normal University.
Consent to participate: Informed consent was obtained from all individual participants included in the study.
Consent for publication: Not applicable.

![]()

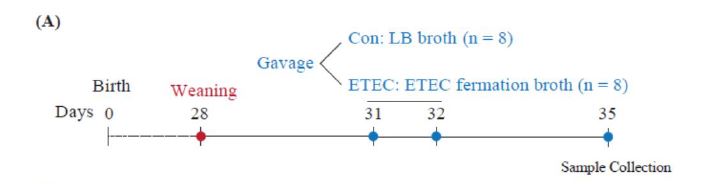 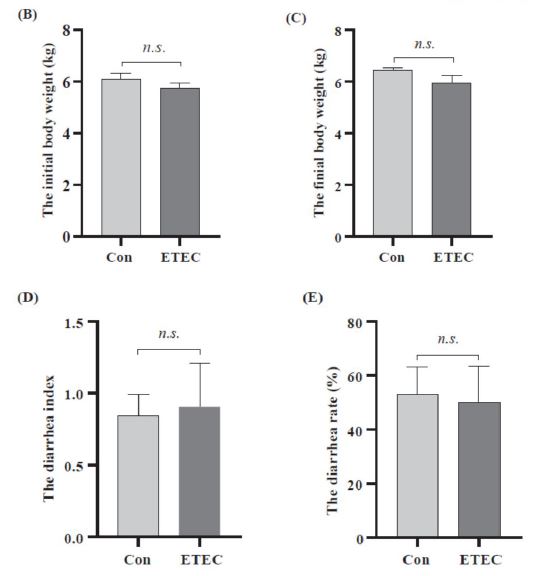 |
| Figure 1: Effect of ETEC challenge on growth performance and fecal score. (A) Experimental outline in Ningxiang piglets. (B) The initial body weight at 21 days. (C) The final body weight at 27 days. (D-E) The diarrhea index and diarrhea rate after ETEC gavage. Values of the bars stand for significant differences using two tailed unpaired t-tests at P < 0.05. n.s., not significant differences. |
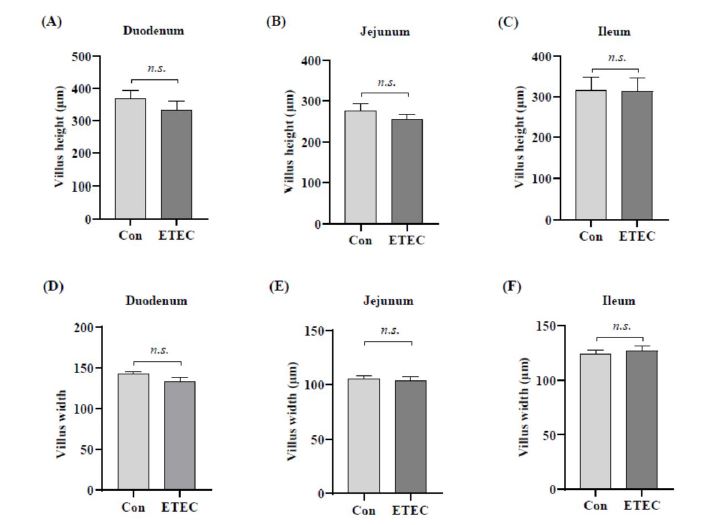 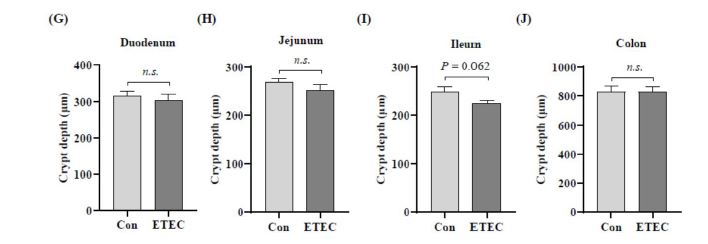 |
| Figure 2: Effect of ETEC challenge on the intestinal morphology of (A-C) villus height in small intestinal, (D-F) villus width in small intestinal, (G-J) crypt depth in small intestinal and in colon. Values of the bars stand for significant differences using two tailed unpaired ttests at P < 0.05. n.s., not significant differences. |
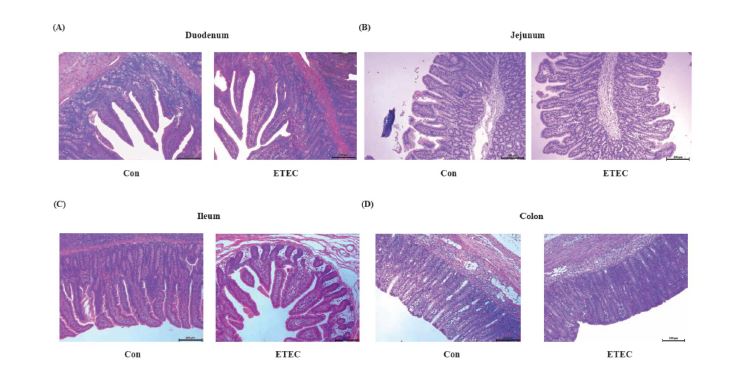 |
| Figure 3: Representative microphotographs of hematoxylin-eosin stained intestinal sections in (A) duodenum, (B) jejunum, (C) ileum and (D) colon. Left: control group; Right: ETEC group. Scale bars, 200 μm. |
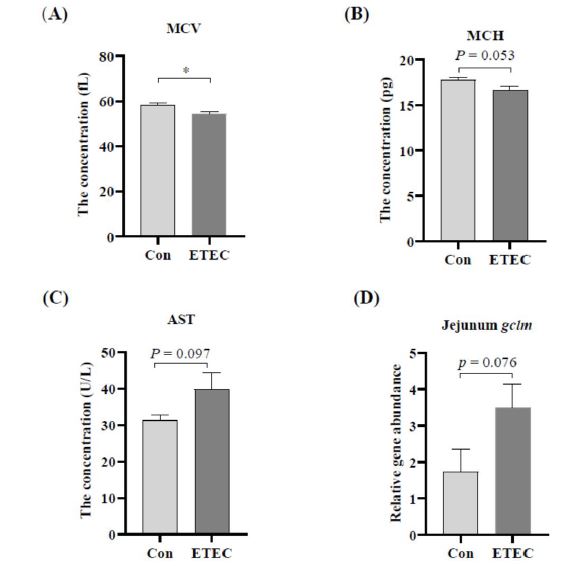 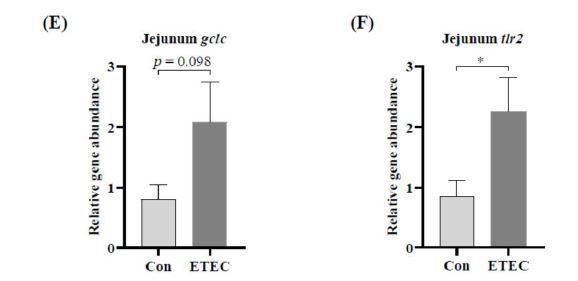 |
| Figure 4: Effect of ETEC challenge in Ningxiang piglets on the levels of (A) MCV in blood, (B) MCH in blood and (C) AST in serum. (D-F) At the transcriptional level, gene expression differences in relative to β-actin analyzed by real-time PCR. Values of the bars stand for significant differences using two tailed unpaired t-tests at P < 0.05. n.s., not significant differences. |
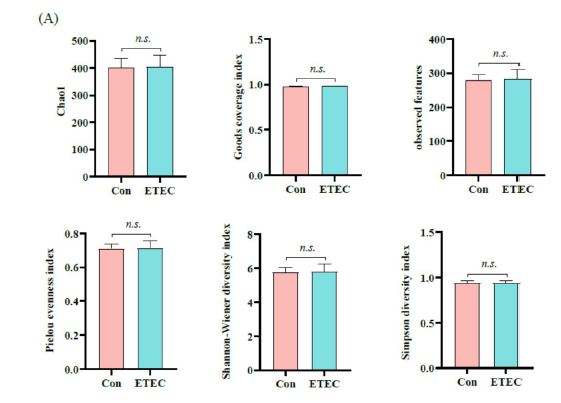 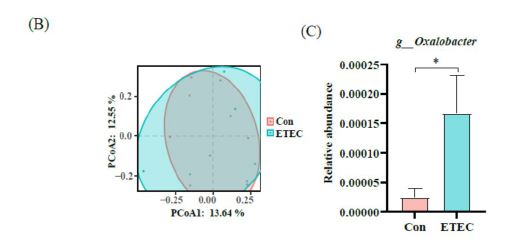 |
| Figure 5: The 16S rRNA gene sequencing analysis of ETEC challenge in Ningxiang piglets on colonic microbiome. (A) Alpha diversity comparisons of the gut microbiome, including Chao1, Goods coverage index, observed features, Pielou evenness index, Shannon-Wiener diversity index and Simpson diversity index. (B) Scatterplots of PCoA analysis depicting differences in the bacterial community structure on genus level between the control group and ETEC group. (C) The relative abundance of genera with or without ETEC challenge. Values of the bars stand for significant differences using two tailed unpaired t-tests at P < 0.05. |
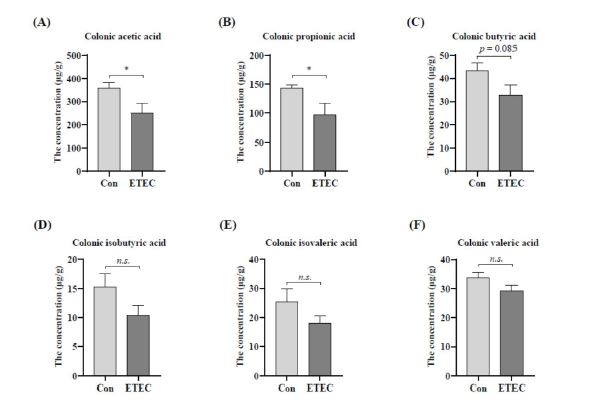 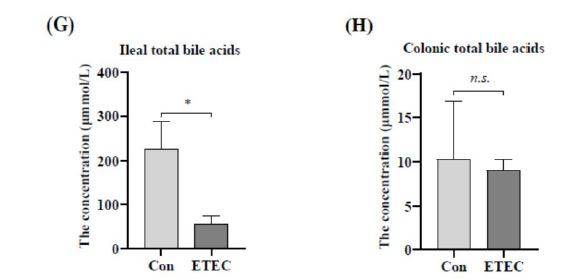 |
| Figure 6: Effect of ETEC challenge on the concentrations of intestinal metabolites, including (A-F) short chain fatty acids in colon, (G) ileal total bile acids and (H) colonic total bile acids. Values of the bars stand for significant differences using two tailed unpaired t-tests at P < 0.05. n.s., not significant differences. |




































