Open Access
Review Article
Max Screen
ISSN: 2348-9790
Copyright: © 2023 Brinderjeet Kaur. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Several scientists called for a new paradigm in animal nutrition by implementing feed diversity in otherwise homogeneously fed livestock production systems as former was shown to have a notable impact on animal productivity and health and the ecological footprint thereby decrease concerns regarding global food security from a societal perspective. However, there are no specific definitions and no threshold values available, so far, for desired grazed grassland feed diversity levels on pastures that enable envisaged sustainable ruminant livestock production and possibly the production of animal products with particular human health-values. Therefore, we conceptualize grazed grassland feed diversity in multiple layers for the very first time in this review to define its relevance from an animal perspective and implications for human health. The data reviewed reveals a close relationship between grazed grassland feed diversity with regard to plant secondary metabolites and animal characteristics that include animal health, welfare, and behavior as well as product quality with possible human health value. Nevertheless, it remains an open question whether plant species diversity per se or increasing occurrence of plant metabolites are responsible for feed diversity effects at grazing. Further, we demonstrate that solely the information about the level of feed diversity present on a field is not sufficient for deducing possible effects on animal related parameters and product quality as other interacting effects such as production level, pedo-climatic effects, and animal breed, have a relevant impact on the actual feed diversity consumed by the animal. Therefore, further research is required to shed light in the dark to understand the interaction between botanical compositions, pasture feed diversity and animal related factors for obtaining benefits for animal health, product quality and its human health value.
Keywords: Secondary Plant Metabolites; Pasture; Feed Biodiversity, Selective Foraging
Cattle livestock production systems in Europe rely on a large quantity of forage and feed produced on arable land and supplied to the animal as mixed rations in confined systems (Schingoethe, 2017; Joubran et al. 2021) [1,2]. In this context, the increasing usage of e.g., corn silage as energy and roughage supplier is of great importance, while in grassland-based systems milk and meat production is dominantly supported by grassland grown feed, offered in mixed rations in confined systems or on pastures. Pastures can provide feed choices and forage diversity for grazing livestock which are crucial for ruminant species to fulfil their physiological demand and express their explorative and selective behavior (Manteca et al. 2008; Villalba et al. 2014; Beck and Gregorini, 2020) [3,4,5]. Nevertheless, entirely pasture-based milk production is of minor importance among the major milk producers in temperate climate, apart from Ireland and New Zealand (Joubran et al.2021) [2]. Recently, however, concerns regarding total mixed ratio (TMR) feeding are arising as this prevents ruminants from displaying their species-specific selective behavior and might, thus, reduce animal welfare (Villalba 2010; Leiber et al. 2020) [6,7]. Grassland-based production comes along with a number of essential services affecting various areas of ruminant livestock production (Peyraud, 2017) [8].
For instance, due to the increased public concern about the potential development of antibiotic resistance in livestock farming, alternatives are being sought which exert a similar prophylactic or therapeutic effect on food-producing animals as conventional treatments in the most natural way (Papatsiros et al. 2012) [9]. Natural substances like plant secondary metabolites (PSM), among others, play an increasingly important role in this respect (Lillehoj et al. 2018) [10]. In addition, some PSM potentially reduce the emission of greenhouse gases by reduction of enteric methanogenesis (e.g., Staerfl et al. 2010; Patra and Saxena 2010) [11,12] or by improvement in internal nitrogen utilization (Kapp-Bitter et al. 2021) [13]. Besides, grazing or grass-based diets enable farmers to meet consumer expectations of livestock systems providing high animal welfare standards (Joubran et al. 2021) [2]. For example, meat or milk from animal-friendly pasture-based production can enable a clear distinction from common agricultural cattle products on the highly competitive global market, thereby generating increased farm income (Kühl et al. 2020; Stampa et al. 2020) [14,15]. Moreover, several studies reported ruminants in grassland-based systems to contribute to food security and wellbeing of communities in grassland areas (Neely et al. 2009) [16]. In fact, when primarily fed with roughage ruminants can reduce competition between livestock feed and human food production as they are able to use human inedible plant resources for producing high nutritional products (Peyraud 2017, Wilkinson and Lee 2018) [8,17]. In addition, soils under permanent grassland contribute notably to carbon sequestration (O´Mara, 2012, Soussana et al. 2014) [18,19]. Increase of grassland and pasture intake in milk production may, therefore, serve as a climate-friendly pathway when compared to confined systems (Lorenz et al. 2019) [20]. However, beside these various potential advantages offered by grassland, the widespread highly intensive dairy systems require diets with high nutrient density what might be a challenging factor. Although complex pasture mixtures for grazing livestock are currently debated as an option to improve animal performance (e.g., Soder et al. 2006; Grace et al. 2018) [21,22], the relationship between animal well-being or performance and feed diversity on grazed permanent grassland has received little attention – especially for dairy cows.
We hypothesize that increasing the feed diversity on grazed permanent grassland could have a beneficial effect on animal health and welfare and animal product quality including its human health value. Therefore, we first try to define, to the best of our knowledge, for the very first time, grassland feed diversity from an animal perspective as there are no threshold values or specific definitions available in literature in this regard (Figure 1). Further, we review the impact of grassland feed diversity with regard to PSM on the health and wellbeing of ruminant livestock species. Moreover, we expand the concept of feed diversity to animal product quality and deduce possible effects on product human health value. Finally, we summarize current literature regarding the inclusion of PSM in ruminant livestock nutrition for potentially mitigating greenhouse gas emissions. Our explanations and considered references are restricted to the temperate grassland regions.
Several scientists called for a new paradigm in animal nutrition by implementing feed diversity as a measure to improve animal wellbeing in otherwise homogeneously fed livestock production systems (Leiber et al. 2020) [7]. Irrespectively of feed provision on pasture or in housing, a mixed diet consisting of a range of individual items in varying physico-chemical constitution offering the animal to deliberately chose is regarded as diverse feed (Gregorini et al. 2017) [23]. However, there is i) no strict definition of feed diversity from an animal perspective - neither for pasture nor for confined systems, ii) no information available to what extent or which component of feed diversity is actually responsible for particular animal effects, iii) no threshold value available for critical feed diversity levels needed under varying nutritional status, health condition, type and breed of livestock or production purpose (wool, milk, meat) to reach desired outcomes and iv) no information available on whether feed diversity on pasture causes additional effects compared to identical amounts fed indoors. In addition, at least for grazing, it is not clear to which functional unit feed diversity refers to other than to offer structurally, chemically, and functionally diverse feed resources (Provenza et al. 2007; Gregorini et al. 2017) [23,24].
Intuitively speaking, the diversity of plant species, functional groups or varieties of a plant species would be conceivable in terms of feed diversity on pasture. Variation of these individual components of plant diversity may increase the overall feed diversity offered and also animal benefits if the identity of available species supports livestock (Distel et al. 2020) [25]. Monocotyledonous (grasses) and dicotyledonous species (legumes, herbs) differ in their chemical constitution in terms of nitrogen concentration (Whitehead 2000) [26], carbohydrate fractions (Gierus et al. 2012) [27] or PSM (Mueller-Harvey 2006) [28], the rate of change of these nutrients (Lüscher et al. 2014) [29], the time of peak nutrient concentration and their overall rate of maturation (Calvière and Duru 1995) [30]. However, there is a difference between the level of feed diversity offered on pasture and that actually consumed as consequence of animal related effects such as preference (Parsons et al. 1994) [31], selection (Hodgson 1990) [32] or a trade-off in benefits against cost of obtaining feed diversity in a meal on pasture as consequence of spatial segregation (Dumont et al. 2002; Koczura et al. 2019) [33,34] and plant related effects like odor, taste (Villalba et al. 2014) [4] and phenological stage (Dumont et al. 1995) [35]. The aforementioned attributes likely explain differences in pasture intake rate between plant species (Rook et al. 2002; Pembleton et al. 2016) [36,37] and consequently the level of feed diversity actually realized.
Balancing between the attributes inherent to grazing livestock to approach the forage source, the amount of herbage on offer and its actual physico-chemical constitution may help if it is the aim to assess principles between pasture feed diversity and grazing livestock responses. It may be important to distinguish between the present feed diversity on pasture and the level utilized by livestock since, at least for dairy cows, thorny or toxic species or those that are senescent are usually avoided. Hence, feed diversity may be a more appropriate term than phytodiversity in its botanical sense when taking the animal perspective into consideration.
The amount of daily herbage consumed by grazing livestock is the product of the time spent grazing and the rate of herbage intake during a grazing event. It is the product of the biting rate and the herbage intake per individual bite (bite mass) (Hodgson 1990) [32]. The functional unit describing grazing behavior consequently is the bite, its mass and the rate of bites (Spedding et al. 1966) [38]. Each bite represents a feed item. The feed item is then a functional unit describing actual feed diversity realized in the ingested herbage as a portion of the total feed diversity available. The actual feed diversity, consequently, may deviate from the level of diversity offered, which may be termed potential feed diversity. Latter is the level offered on pasture in terms of botanical and physico-chemical diversity in space and time. The level of actual feed diversity is limited by the ability of animals in a way that the amount realized depends on several interacting factors (Figure 1). Limitations in the realizable feed diversity are either driven by exogeneous constraints such as pasture access, variability of feeding stations/patches and its distribution or by endogenous animal factors of the grazing species. For instance, the herbage intake is closely related to the animal size, its potential productiveness (Holmes 1989) [39] or stage of development (Pembleton et al. 2016) [37] when herbage availability in amount and quality is adequate. Animals not only select different plant species, but also different plant organs (Bullock and Marriott 2000) [40] although sheep and cattle differ in their anatomical ability to select for particular plant parts. Sheep nibble and bite rather than tearing the herbage biomass as is typical for cattle (Holmes 1989) [39]. These factors determine that actual feed diversity is lower than the potential (Figure 1). Differences in the level of feed diversity offered and realized by the grazer are likely when considering that dairy cows ingest organic matter of higher digestibility than is on average offered per pasture (Dodd et al. 2018) [41].
When feed items offered against selected ones are structured according to single bites, the feed diversity realized in single bites may be determined and seen as α-diversity. Such information may, for instance, be obtained by continuous bite monitoring according to the method described by Bonnet et al. (2015) [42]. The feed diversity realized by the grazing livestock is likely related to the diversity offered per feeding station (α-diversity), between feeding stations (β-diversity), the total pasture scale-diversity (γ-diversity) and their spatial arrangement. Metabarcoding may help to determine the realized feed diversity in diets based on chloroplast DNA obtained from feces (Palumbo et al. 2021) [43]. It is also likely that the forage quality or specific metabolites present in plants within patches reinforce the relationship between offered and realized feed diversity (Figure 1) and the pasture-scale forage quality changes over time and due to stocking intensity (Pavlu et al. 2021) [44]. Moreover, although the α-diversity per bite is likely bigger for cattle than for sheep, this must not be valid for the actual α-diversity per kg LW0.75 nor the β-diversity when considering all bites in one meal. It is, however, not clear whether realized feed diversity in herbage intake is the result of potential feed diversity offered or the cause. Attempts have been made in earlier studies in a way to describe the offered and ingested herbage in terms of phenological stage and botanical composition for beef (e.g., Dumont et al. 2007) [45] and dairy cattle (Farruggia et al. 2014; Koczura et al. 2019) [34,46]. These studies, however, did not evaluate feed diversity offered and utilized by livestock from an animal perspective, so far. Comparing levels of realized feed-diversity (α- or β-diversity) to the potential under varying environmental conditions or livestock production purpose and considering a range of desired outcome variables (livestock productivity, wellbeing, or health) on a range of livestock species could shed light in the dark to understand the role of feed diversity for animals.
Several aspects of potential feed diversity in terms of functional traits, functional groups, varieties, or species richness have received recognition in studies with focus on herbage quality (Komainda and Isselstein 2020) [47], livestock performance (Grace et al. 2019) [48], PSM and enteric methane emission (Carmona-Flores et al. 2020; Loza et al. 2021) [49,50] or productivity (Lorenz et al. 2020) [51]. Pastures for dairy cows are usually high in herbage organic matter digestibility and they are grazed in daily portions or rotationally at defined herbage allowances under high stocking density (Delaby et al. 2020) [52]. The pastures are thus managed in a way to increase spatial homogeneity of forage items and their quality. Feed diversity under such circumstances refers then to the diversity of different plant species, functional groups or plant functional types (Andueza et al. 2010) [53] and their evenness in the sward rather than to the diversity of plant species in different phenological stages as is common for low-input grassland (Pavlu et al. 2006) [54]. However, beneficial effects on animal health or production also depend on the species identity and whether or not attributes present in these species are supportive (e.g., PSM such as condensed tannins in some legumes, iridoid-glycosides, or polyphenols in herbs) (Bryant et al. 2018; Kapp-Bitter et al. 2021) [13,55]. For instance, Kapp-Bitter et al. (2021) [13] tested 35 different hays produced from plant species containing PSM on rumination characteristics in vitro and found a relevant potential improvement in nitrogen utilization. Woodward et al. (2001) [56] found that ruminants fed diets based on Lotus spp. that contain PSM have lower methane emission per unit dry matter intake. However, such PSM-rich species are obviously not ingested in sufficient amounts at grazing (Loza et al. 2021) [50]. Is the lack of low amounts of PSM-containing plant species in the ingested herbage only due to limited yield proportions and an insufficient distribution or is the motivation of the animals to avoid certain plant species expressed here, so that the question arises under which conditions cows ingest larger amounts of PSM-containing species during grazing in order to achieve certain objectives. Understanding realized feed diversity in relation to the distribution of distinct species in swards is critical to the design of future environmental-friendly livestock production systems. The crude protein concentration may serve as indicator of realized feed diversity in dairy cow pastures since it is negatively correlated to the herbage accumulation and phenological development (Américo et al. 2021) [57]. Legumes rich in PSM usually also contain high concentrations of crude protein and associative effects on herbage intake in mixed swards may result when partner species have lower protein concentration (i.e., old grass) (Niderkorn and Baumont 2009) [58]. It must be assumed that relationships are not linear, because mutualistic or synergistic relationships exist between plant species in mixed swards, which is expressed, for example, in a spatial variation of the protein concentration of grasses depending on the neighbouring species (legume) (Opitz von Boberfeld and Biskupek, 1995) [59]. Farruggia et al. (2006) [60] and Koczura et al. (2019) [34] found clear preferences for vegetative grasses with higher digestibility on diverse mountain pastures when these were easily accessible for dairy cows. With advances in maturity, the selectivity for grasses progressively declined in that latter study but this also coincided with the lactation stage. Grazing management that presents cows desired feed diversity and meets dietary preferences will be the challenge to manage biodiversity of grassland and to use biodiversity for management of dairy cows.
To evaluate the relationship between grassland phytodiversity and dairy cow performance a scientific literature search was conducted using the online databases Web of Science and Google Scholar. The search used included “dairy cow performance OR milk yield OR milk production OR dairy cow milk yield”, “grazing OR pasture”, “multispecies OR complex sward OR plantain OR herb inclusion”. For the purpose of this review, peer-reviewed literature was considered as the minimum reporting standard required for inclusion. Reference lists from papers were reviewed for any additional studies relevant to the review and only studies from the last 20 years were searched in general. Eligible studies were included for further consideration if (i) the study was reported in English; (ii) the population in the study was grazing dairy cows; and (iii) the treatment of interest included at least one simple reference sward (e.g. white clover-grass) in comparison to at least one complex multispecies sward (e.g. white clover-grass plus herbs) offered through grazing in temperate agroclimate. Studies with inadequate methodologies or reporting were excluded. Results in studies reported over several periods were treated as separate experiments unless response variables (e.g. milk production) were averaged over periods within the results. In those cases, average values were obtained across periods (e.g. Soder et al. 2006) [21]. Interest was given to the relationship of dairy cow performance expressed as energy-corrected milk (ECM, kg cow-1 day-1) calculated in accordance with (Brade and Flachowsky 2005) [61] which expresses the milk production corrected for protein and fat (4% fat, 3.4 % protein). Results given as milk volume (L cow-1 day-1) were transformed into milk mass (kg cow-1 day-1) according to O’Connor (1995) [62]. In total 16 studies were considered suitable (Table 1) and resulted in 60 treatment combinations for comparisons of simple reference swards against complex multispecies swards. The reference swards were not necessarily pure stands of e.g. grasses but less complex in terms of species richness than the complex sward treatment that was used in the respective comparison (Table 1).
Among the studies reviewed, diversification in grazed grassland for dairy cows mainly refers to the cultivation of sown multispecies sward with more or less complexity (Table 1). In the studies reported in Table 1, all swards consisted of pre-selected species established shortly before the experiments and only 18 plant species were sown in total (Table 1). This is only a fraction considering that dairy cows on Italian summer pastures in the Alps ingested up to 39 plant species (Palumbo et al. 2021) [43].
The botanical composition and its physico-chemical structure are usually shaped over time by defoliation intensity, edaphic factors, and pedo-climatic conditions (Michaud et al. 2012) [63] and therefore potential feed diversity is likely amplified on permanent grassland. Strictly speaking, the conditions and comparisons of milk response to feed diversity, so far, do not refer to permanent grassland (Peeters et al. 2014) [64]. The few exceptions with comparative studies of milk response on permanent grassland were located in France (Farruggia et al. 2014; Koczura et al. 2019) [34,46] and they may be considered outliers in the dataset since the botanical composition differs from the studies using sown swards. Of the 60 study comparisons reviewed, 25 were found with a negative response in dairy cow performance and 35 cases where the response was positive (Table 1). To test for the milk response statistically analysis of covariance was used applying a linear-mixed effects model with the categorical factors i) sward treatment (complex vs. reference) and ii) lactation stage (early, mid, late) and iii) the continuous variable organic matter digestibility (OMD, %) as fixed and interactive predictor effects in the R package ‘nlme’. The experiment was a random effect. After automatic model reduction using the 'MuMIn' package, the two-fold interaction between lactation stage and sward treatment (F=4.5, p=0.036) was significant for the dairy cow performance. Posthoc comparison of means using the Tukey’s test in the ‘emmeans’ package showed that the complex swards resulted in greater dairy cow performance than the references during the early lactation stage (21.7 ± 1.3 vs. 22.8 ± 1.3 kg ECM cow-1 day-1, estimated means ± SE). This difference between swards compares well to the mean increase of daily milk production of 1.2 kg in dairy cows that was found in the meta-analysis of McCarthy et al. (2020) who compiled feeding trials and grazing studies that looked at the influence of inclusion of herb species. In the studies reviewed here, differences between sward treatments during mid and late lactation were, however, not significant (22.2 ± 1.0 and 17.9 ± 1.3 in mid and late lactation, respectively). Some effects of higher pasture diversity may be attributed to identity effects of particular species (Pembleton et al. 2015) [65]. For instance, Pembleton et al. (2016) [37] found a higher milk performance of dairy cows because of inclusion of plantain (Plantago lanceolata L.) while others observed positive responses when chicory (Cichorium intybus) was added (e.g., Roca-Fernandéz et al. 2016) [66]. Under extensive defoliation management and low nutrient input, permanent grassland may develop towards assemblages with high value for biodiversity (Isselstein 2018) [67]. However, to maintain biodiversity in such swards, plants rely on reproductive success which restricts defoliation frequency and, thus, does not permit grazing at phenological stages optimal for dairy cows (Bruinenberg et al. 2002) [68]. A generally positive response to phytodiversity in dairy cow performance will therefore only apply marginally as was noticed by Tonn et al. (2021) [69]. To gain insights into plant species-specific dynamics in forage quality and, hence, to determine the feed diversity in a physico-chemical sense, evaluations at species-level in mixed swards are required which are, so far, extremely rare in grazing studies (e.g. Roca-Fernandez et al. 2016; Loza et al. 2021) [50,66]. This would allow to calculate indices of forage quality diversity, relate these to plant species diversity and also to dairy cow traits. Relating these measures to bites obtained during grazing, would help to evaluate whether plant species richness or physico-chemical richness are more important from an animal perspective.
In their natural feeding behavior, ruminants select and combine forage plants with different nutrient profiles and bioactive substances in their diet (Rutter 2006; Villabla et al. 2013) [70,71] for improving both digestive efficiency and metabolic health (Leiber et al. 2020) [7]. Moreover, there is scientific evidence that selectivity of forage items is an important attribute to perform the intrinsic behavior of selecting specific plants which connect physiological needs with sensorial experience and action (Rutter 2006) [70]. Both searching for as well as harvesting the feed belong to them, what also enables animal individual ways of foraging with own choices and decisions (Baumont et al. 2000) [72]. In order to understand the principles of foraging decisions by ruminants two approaches provide contrary explanations. While the synthetic approach assumes that animals behave following an objective, namely, to maximize their ‘fitness’ (Krebs and McCleery 1984; Baumont et al. 2000) [72,73], the analytical approach indicates feed choices are caused by taste (sensory factors) and experiences with pre-choices in diet (Baumont et al. 2000) [72]. These post-ingestive experiences are of greater importance for food choices than solely the sensory factors, thus, food choices can change despite similar food condition (Rook and Tallowin 2003) [74].
It is already well known, if ruminants have the opportunity to select between various plants, they show obvious preference differences e.g., given in intake profiles depending on time of the day. Various studies investigated a characteristic diurnal pattern with greater intake rates for clover during the morning and increasing intake of grass towards the evening for dairy heifers (Rutter et al. 2004a) [75] and dairy cows (Rutter et al. 2004b) [76], respectively. Such behavior was described in Provenza et al. (2007) [24] as well whereby hungry sheep initially preferred digestible instead of less digestible roughage. As sheep continued to graze, they experienced mild aversion against digestible feed items caused by the excess of organic acid due to highly soluble sugars and excess of ammonia from readily degradable proteins and concentrations of toxic cyanogenic glycosides present in legumes. Therefore, they started to ingest a higher amount of grass in order to reduce fermentation rate and increase ruminal buffer capacity (Provenza et al. 2007) [24]. Considering this variation in feed choices and diet selection in cattle, the limitation of this feeding behavior through pastures consisting of only one plant species could lead to stress and impairments in animal welfare (Manteca et al. 2008) [3]. Most species-poor grasslands provide only minor or no option to express the above-mentioned diurnal pattern in diet selection in cattle, what could come along with frustration on the animal side when experienced over longer periods since herbage intake takes a significant portion of the time on pastures (Rutter 2006) [70]. In this respect, Beck and Gregorini (2020) [5] stated, wellbeing in livestock could be reached when they are able to pursue their full potential to express behavior. However, the effects of grassland feed diversity on dairy cattle behavior (except feeding behavior) are only minor reported in literature, to the authors knowledge. For heifers, Hejcmanová et al. (2009) [77] described, that animals under an intensive grazing treatment on species-rich upland pasture showed less time ruminating and resting compared to those in the extensive treatment. The assessment of species-rich permanent grassland and pasture in terms of dairy cattle behavior traits of e.g., various breeds need further investigations.
As mentioned earlier, taste, smell and sight are labels which animals use to select feed (Villalba 2010) [6] and associate these sensorial factors with the bringing post-ingestive feedback. Thus, animals learn to use the medical properties of bioactive secondary compounds in plants to enhance nutritional and health status when exposed to variable diets (Villalba et al. 2014) [4].
Many studies revealed a medicinal potential of PSM against gastrointestinal parasites (Hoste et al. 2006; Villalba et al. 2014) [4,78]. However, there is the necessity to further investigate the mechanisms of action and the synergistic relationship among plant metabolites in order to increase efficiency of phytotherapy for control of gastrointestinal parasites in ruminants as for many bioactive compounds in vitro tests revealed promising results in contrast to in vivo attempts (Hoste et al. 2006; Ku-Vera et al. 2020) [78,79]. Further, condensed tannins as present, for instance, in some forage legumes (e.g., Lotus corniculatus, Onobrychis viciifolia) were shown to diminish the risk for pasture bloat in cattle (Rochfort et al. 2008) [80].
Many antiparasitic and antioxidant secondary compounds have a bitter and adverse taste (Villalba et al. 2014) [4]. However, it is known that herbivores change feeding behavior in response to parasitic infection. For instance, sheep parasitized with Haemonchus contortus were shown to ingest more tannin-rich plants than non-infected control groups (Lisonbee et al. 2009) [81]. Likewise, Amit et al. (2013) [82] observed a higher ingestion of tannin-rich feed of infected Mamber goats compared to their non-infected counterparts when exposed to a free choice between tannin-free hay and Pistacia lentiscus leaves high in condensed tannins. Also, Amit et al. (2013) [82] reported breed differences for self-medication as infected Damascus goats did not show a higher intake of Pistacia lentiscus, explaining such behavior with the fact that latter is consuming the plant already before being infected whereas Mamber goats use the plant for curative self-medication. In contrast, Gaudin et al. (2018) [83] observed no significant differences between intake of tannin-rich Onobrychis viciifolia compared to tannin-free Medicago sativa in infected and non-infected kids and lambs, which the authors explain by the fact that forage with high energy and protein concentrations may help animals to control parasitic infection without the urgency to ingest tannin-rich feed resources. In fact, Houdijk (2012) [84] reviewed that host resistance against gastrointestinal nematodes can be improved by supplementing protein rich foods which enhance the nutritional status as well as the immune status of the host. Further, Hutchings et al. (2002) [85] observed that infected animals avoided grazing near dung pats even when those areas offered higher nutrient rewards. In addition, parasitized animals, when forced to consume contaminated pasture, grazed further from the soil surface than non-infected animals (Hutchings et al. 2003) [86]. Former behavioral patterns could be explained by the already mentioned learning process between orosensorial characteristics of forage items and postingestive feedback (Provenza and Villalba, 2006; Villalba et al. 2014) [4,87]. Finally, prophylactic behavioral patterns in ruminants might be explained by genetic adaptation to a respective environment (Villalba et al. 2014) [4].
Literature reveals that animal products (e.g., milk), which are produced with botanically diverse forage, contain higher amounts of PSM such as terpenes or phenols than the one based on botanically poor feed (Avondo et al. 2013) [88]. In particular, terpenes are known to have an effect on the aromatic profile and on the sensory characteristics of dairy products (e.g., cheese) (Avondo et al. 2013) [88]. Further, terpenes are used as biomarkers to determine pasture feeding and geographical origin in dairy products and meat (Fernandez et al. 2003; Serrano et al. 2011) [89,90]. In addition, PSM (e.g., phenolic compounds, tannins) have been shown to improve color stability, flavor, shelf life and odor in products produced by ruminants (Beck and Gregorini 2020) [5]. In an experiment of Nieto et al. (2011) [91], the inclusion of distilled rosemary leaf in the feeding ratio of pregnant ewes reduced lipid oxidation, odor, and flavor spoilage in lamb meat. Similarly, Di Trana et al. (2015) [92] observed an increase of polyphenols in plasma and in milk of Girgentana goats, when including Sulla fresh forage (Sulla coronarium L.) in the diet, which improved the antioxidant activity in their milk. Likewise, Bonanno et al. (2019) [93] increased the efficiency of the immune system of dairy cows and improved the oxidative stability and antioxidant properties of cheese when including wheat bran in the diet. Similarly, Luo et al. (2019) [94] measured higher radical-scavenging ability and antioxidant capacity in lamb meat fed on pasture grass compared to lamb meat fed on mixed diets.
Next to improved product quality characteristics, PSM are known to reduce bio-hydrogenation in the rumen and, therefore, facilitate the transfer of polyunsaturated fatty acids (PUFA) in dairy products and meat (Vasta and Luciano 2011; Schulz et al. 2018) [95,96]. Higher contents of unsaturated fatty acids, especially n-3 PUFA, are known to reduce the risk for cardiovascular diseases and high blood pressure (Li et al. 2015; Mozaffarian et al. 2010) [97,98]. In fact, when replacing 5% of SFA with PUFA it was shown that the risk for cardiovascular diseases could be reduced by 17-25% (Li et al. 2015; Mozaffarian et al. 2010) [97,98]. Several studies could show that feeding botanical diverse feed resources in form of pasture improves the fatty acid profile in livestock products. For example, O’ Callaghan et al. (2016) [99] illustrated in their real-time comparison of distinct feeding systems that pasture use, overall, had a positive effect on milk quality (fat, protein) and further increased concentrations of PUFA and n-3 FA compared to milk produced in TMR feeding systems. Likewise, Hofstetter et al. (2014) [100] observed significantly higher concentrations of PUFA and n-3 FA in the pasture feeding system than in the indoor feeding system. In Schulz et al. (2018) [96] an increase in PUFA and a reduction in the n-6/ n-3-ratio was measured when replacing maize silage with red clover (Trifolium pratense) silage, as protein-bound phenol formations, triggered by the enzyme polyphenol oxidase present in red clover protects dietary PUFA against biohydrogenation. Lastly, French et al. (2000) [101] demonstrated that PUFA can be increased in the intramuscular fat from steers when including grazed grass in the diet.
Extensively managed biodiversity-oriented pastures, characterized by continuous grazing with low stocking rate, as shown in Farruggia et al. (2014) [46], exhibited a greater Shannon diversity of flower visiting insects (e.g. Lepidoptera sp.) than intensive production oriented pastures, characterized by rotational grazing and higher stocking rate. In terms of sward productivity, however, plant species diversity had only a positive impact under an ungrazed cutting regime (Jing et al., 2017) [102]. Moreover, including diverse feed resources rich in PSM were described to reduce the emission of greenhouse gases like methane and possibly nitrogen excretion into the environment (Gardiner et al. 2016; Ku-Vera et al. 2020) [79,103]. For instance, Staerfl et al. (2010) [11] revealed, with the in vitro Hohenheim gas test, that the inclusion of polyphenols in cows’ diets decreased methane emissions as polyphenols optimize rumen fermentation and reduce protozoa associated with methanogenesis (Staerfl et al. 2010) [11]. Likewise, Patra and Saxena (2010) [12] stated in their review, that saponins and tannins, present in a wide variety of plants, may decrease methanogenesis in the rumen directly through reducing activity of methanogens and indirectly through inhibition of rumen protozoal growth. In addition, polyphenols and tannins in ruminants’ diets are reported to reduce protein degradation in the rumen and consequently increase amino acid availability for intestinal adsorption and reduce nitrogen excretions into the environment (Avondo et al. 2013; Lillehoj et al. 2018) [10,88]. Any measure that helps to minimize systemic carbon and nitrogen emissions that inevitably arise from dairy production, therefore, makes a significant contribution to protect the climate in the long term. Targeted use of plants with desirable secondary ingredients could, consequently, be a key factor and part of that may be to grow multispecies swards which need less fertilizer input and produce less greenhouse gases (Cummins et al. 2021) [104]. Lastly, Zytynska and Meyer (2019) [105] reviewed that high plant species richness has a stabilizing effect on multitrophic interactions like rates of herbivory and predation in grassland ecosystems and therefore might enhance resilience of grassland in terms of pests [106-114].
The review study aimed to provide a holistic approach in describing the effect of grassland feed diversity on animal food production in order to follow the growing demand for a new paradigm in ruminant livestock nutrition by implementing feed diversity from an animal perspective for improving animal wellbeing.
In conclusion, we provide evidence for a close relationship between feed diversity of grassland with special regard to PSM and animal characteristics that include product quality and health of ruminants. In addition, we described that the inclusion of plants rich in PSM might have positive effects on product human health value. Further, feed diversity of grassland may have a direct beneficial emission in livestock production on the environment. Moreover, we demonstrate that solely the information about the level of feed diversity present on a field is not sufficient for deducing possible effects on animal related parameters and product quality as other interacting effects such as production level, pedo-climatic effects, and animal breed, have a relevant impact on the actual feed diversity consumed by the animal. Therefore, there is high uncertainty about the specific mechanistic principles which are underlying between the level of feed diversity offered and the actual feed diversity realized in feed items (bites) by the grazing cow stressing the knowledge gap in milk response to feed diversity on typical permanent grassland. Consequently, further research is required to fully understand the interaction between botanical composition, pasture feed diversity and animal related factors (e.g., breed) for obtaining benefits for animal health, product quality with its human health value and the environment.
Conceptualization, investigation, resources, T.Z., M.K., S.A; writing original draft preparation, funding acquisition T.Z; writing, review-editing T.Z., M.K., S.A; visualization M.K.; project administration, supervision J.I. and M.G.
Authors declare no conflict of interest
This work was supported by the Open Access Publishing Fund provided by the Free University of Bozen-Bolzano.
This work was supported by the Open Access Publishing Fund of the Free University of Bozen-Bolzano.

![]()

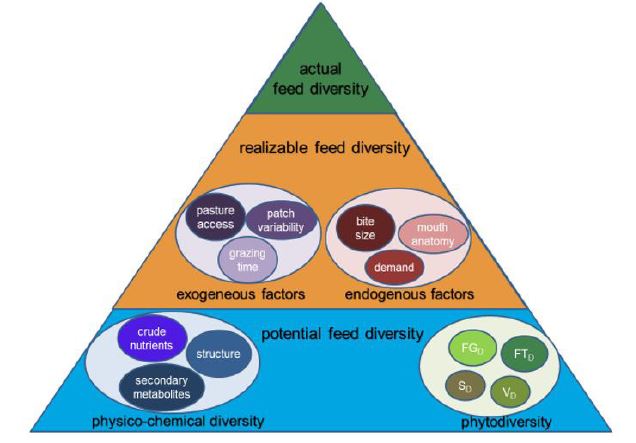 |
| Figure 1: Conceptualization of feed diversity in multiple layers from potential feed diversity offered to that one realizable by the animal and the actual feed diversity which is realized FGD: Functional Group Diversity; FTD: Functional Trait Diversity; SD: Species Diversity, VD: Variety Diversity |
Reference |
DIM |
complex sward treatment |
ECM complex |
OMD complex |
simple sward treatment |
ECM simple |
OMD simple |
ECM difference |
Box et al. 2017 |
217 | PRG-WC | 19.3 | 62.7 | PL pure | 21.5 | 67.0 | -2.2 |
Box et al. 2017 |
217 | PRG-WC-PL | 20.7 | 64.9 | PL pure | 21.5 | 67.0 | -0.7 |
Box et al. 2017 |
51 | PRG-WC | 33.4 | 63.9 | PL pure | 32.7 | 65.3 | 0.6 |
Box et al. 2017 |
51 | PRG-WC-PL | 33.7 | 65.2 | PL pure | 32.7 | 65.3 | 0.9 |
Bryant et al. 2017 |
97 | PRG 2n-WC-Bromus-Chic-PL-RC | 23.1 | 73.9 | PRG 2n-WC | 22.1 | 74.4 | 1.0 |
Bryant et al. 2017 |
97 | PRG HSG-WC-Chic-PL-Lot ped | 22.5 | 76.2 | PRG HSG-WC | 22.8 | 76.7 | -0.3 |
Bryant et al. 2017 |
97 | Fear-WC-Bromus-Chic-PL-LUC | 21.9 | 70.0 | FA-WC | 24.5 | 77.3 | -2.6 |
Bryant et al. 2017 |
154 | PRG 2n-WC-Bromus-Chic-PL-RC | 21.9 | 69.9 | PRG 2n-WC | 18.8 | 74.0 | -1.3 |
Bryant et al. 2017 |
154 | PRG HSG-WC-Chic-PL-Lot ped | 19.0 | 75.5 | PRG HSG-WC | 20.3 | 78.8 | -2.7 |
Bryant et al. 2017 |
154 | Fear-WC-Bromus-Chic-PL-LUC | 19.5 | 68.1 | FA-WC | 22.2 | 74.3 | 0.3 |
Bryant et al. 2017 |
237 | PRG 2n-WC-Bromus-Chic-PL-RC | 18.1 | 78.8 | PRG 2n-WC | 17.9 | 79.1 | 0.3 |
Bryant et al. 2017 |
237 | PRG 2n-WC-Bromus-Chic-PL-RC | 18.2 | 82.0 | PRG HSG-WC | 19.4 | 83.1 | -1.2 |
Bryant et al. 2017 |
237 | PRG 2n-WC-Bromus-Chic-PL-RC | 17.7 | 77.4 | FA-WC | 18.8 | 80.4 | -1.0 |
Bryant et al. 2018 |
193 | PRG-WC-Ital RG | 18.6 | 75.1 | PRG-WC | 19.7 | 75.4 | -1.1 |
Bryant et al. 2018 |
193 | PRG-WC-Chic-PL-LUC | 20.9 | 74.8 | PRG-WC | 19.7 | 75.4 | 1.1 |
Bryant et al. 2018 |
193 | PRG-WC-Chic-PL-LUC-Ital RG | 21.7 | 75.5 | PRG-WC | 19.7 | 75.4 | 2.0 |
Carmona-Flores et al. 2020 |
169 | Fear-PRG-WC-PL-Chic-Lot corn mix | 27.4 | 68.4 | PRG-WC mix | 25.0 | 66.9 | 2.5 |
Carmona-Flores et al. 2020 |
169 | Fear-PRG-WC-PL-Chic-Lot corn mix | 30.5 | 68.3 | PRG-WC spatial | 26.5 | 69.5 | 4.0 |
Chapman et al. 2008 |
22 | Ital RG-T michel-WC | 12.9 | 56.9 | PRG-WC-T subter | 10.7 | 64.5 | 2.2 |
Chapman et al. 2008 |
22 | Fear-WC-T subter | 13.1 | 59.6 | PRG-WC-T subter | 10.7 | 64.5 | 2.4 |
Chapman et al. 2008 |
22 | Fear-Dact-WC-RC-Chic | 14.1 | 64.2 | PRG-WC-T subter | 10.7 | 64.5 | 3.4 |
Dodd et al. 2018 |
200 | PRG-LUC-PL | 13.5 | 59.8 | PRG-LUC | 14.5 | 56.6 | -1.0 |
Dodd et al. 2018 |
200 | PRG-LUC-PL | 17.2 | 59.1 | Fear-LUC | 14.3 | 57.8 | 2.8 |
Dodd et al. 2018 |
78 | PRG-LUC-PL | 20.1 | 69.2 | PRG-LUC | 20.7 | 69.3 | -0.6 |
Dodd et al. 2018 |
78 | Fear-LUC-PL | 21.1 | 67.2 | Fear-LUC | 20.5 | 66.9 | 0.7 |
Farruggia et al. 2014 |
131 | diverse PGL | 16.8 | 67.0 | semi-diverse PGL | 17.2 | 75.0 | -0.4 |
Jonker et al. 2019 |
162 | PRG-WC-Ital RG-LUC-Chic-PL | 19.9 | 66.9 | PRG-WC-Ital RG | 18.1 | 66.9 | 1.7 |
Koczura et al. 2019 |
197 | high-biodiv PGL | 16.1 | 59.7 | low-biodiv PGL | 18.8 | 66.6 | -2.7 |
Loza et al. 2021 |
45 | PRG-WC-RC-Chic-PL-SB-CAR-Lot corn | 29.9 | 84.4 | PRG-WC | 28.9 | 87.6 | 1.0 |
Loza et al. 2021 |
154 | PRG-WC-RC-Chic-PL-SB-CAR-Lot corn | 23.3 | 77.9 | PRG-WC | 22.0 | 80.2 | 1.3 |
Nkomboni et al. 2021 |
279 | PRG-WC-PL_15 | 20.6 | 78.7 | PRG-WC | 19.7 | 76.5 | 0.9 |
Nkomboni et al. 2021 |
279 | PRG-WC-PL_30 | 20.4 | 78.1 | PRG-WC | 19.7 | 76.5 | 0.7 |
Nkomboni et al. 2021 |
279 | PRG-WC-PL_60 | 20.6 | 78.3 | PRG-WC | 19.7 | 76.5 | 0.9 |
Pembleton et al. 2016 |
125 | PRG-WC-PL mix dryland | 21.8 | 59.8 | PRG dryland | 22.2 | 60.4 | -0.5 |
Pembleton et al. 2016 |
125 | WC-PL dryland | 19.5 | 51.3 | PRG dryland | 22.2 | 60.4 | -2.7 |
Pembleton et al. 2016 |
125 | PRG-WC-PL spatial dryland | 21.4 | 60.1 | PRG dryland | 22.2 | 60.4 | -0.8 |
Pembleton et al. 2016 |
246 | PRG-WC-PL mix dryland | 17.9 | 70.4 | PRG dryland | 17.7 | 71.4 | 0.2 |
Pembleton et al. 2016 |
246 | WC-PL dryland | 15.4 | 64.9 | PRG dryland | 17.7 | 71.4 | -2.3 |
Pembleton et al. 2016 |
246 | PRG-WC-PL spatial dryland | 17.5 | 67.6 | PRG dryland | 17.7 | 71.4 | -0.3 |
Pembleton et al. 2016 |
35 | PRG-WC-PL mix dryland | 23.9 | 66.1 | PRG dryland | 22.5 | 64.2 | 1.3 |
Pembleton et al. 2016 |
35 | WC-PL dryland | 23.1 | 65.3 | PRG dryland | 22.5 | 64.2 | 0.5 |
Pembleton et al. 2016 |
35 | PRG-WC-PL spatial dryland | 23.7 | 66.7 | PRG dryland | 22.5 | 64.2 | 1.2 |
Pembleton et al. 2016 |
125 | PRG-WC-PL mix irrigated | 21.8 | 67.8 | PRG irrigated | 22.2 | 66.1 | -0.5 |
Pembleton et al. 2016 |
125 | WC-PL irrigated | 19.5 | 60.2 | PRG irrigated | 22.2 | 66.1 | -2.7 |
Pembleton et al. 2016 |
125 | PRG-WC-PL spatial irrigated | 21.4 | 63.9 | PRG irrigated | 22.2 | 66.1 | -0.8 |
Pembleton et al. 2016 |
246 | PRG-WC-PL mix irrigated | 17.9 | 71.8 | PRG irrigated | 17.7 | 74.5 | 0.2 |
Pembleton et al. 2016 |
246 | WC-PL irrigated | 15.4 | 68.0 | PRG irrigated | 17.7 | 74.5 | -2.3 |
Pembleton et al. 2016 |
246 | PRG-WC-PL spatial irrigated | 17.5 | 71.8 | PRG irrigated | 17.7 | 74.5 | -0.3 |
Pembleton et al. 2016 |
35 | PRG-WC-PL mix irrigated | 23.9 | 68.0 | PRG irrigated | 22.5 | 68.9 | 1.3 |
Pembleton et al. 2016 |
35 | WC-PL irrigated | 23.1 | 66.6 | PRG irrigated | 22.5 | 68.9 | 0.5 |
Pembleton et al. 2016 |
35 | PRG-WC-PL spatial irrigated | 23.7 | 67.7 | PRG irrigated | 22.5 | 68.9 | 1.2 |
Roca-Fernandéz et al. 2016 |
181 | PRG-WC-RC | 17.3 | 78.6 | PRG | 16.2 | 78.8 | 1.1 |
Roca-Fernandéz et al. 2016 |
181 | PRG-WC-RC-Chic | 17.7 | 79.8 | PRG | 16.2 | 78.8 | 1.5 |
Roca-Fernandéz et al. 2016 |
181 | PRG-WC-RC-Chic-Fear | 17.9 | 79.4 | PRG | 16.2 | 78.8 | 1.8 |
Soder et al. 2006 |
109 | Dact-WC-Chic | 31.7 | 61.3 | Dact-WC | 30.7 | 58.5 | 1.0 |
Soder et al. 2006 |
109 | Dact-Fear-PRG-RC-Lotus corn-Chic | 31.1 | 58.2 | Dact-WC | 30.7 | 58.5 | 0.4 |
Soder et al. 2006 |
109 | Dact-Fear-PRG-RC-Lot corn-Chic-WC-LUC-Poa prat | 30.3 | 61.8 | Dact-WC | 30.7 | 58.5 | -0.4 |
Totty et al. 2013 |
190 | PRG HSG-WC-Chic-PL-Lotus ped | 20.0 | 78.6 | PRG HSG-WC | 19.6 | 78.4 | 0.4 |
Wilson et al. 2021 |
157 | Festulolium-Fear-Dact-WC | 21.6 | 64.8 | Chic-PL-WC | 24.0 | 65.4 | -2.3 |
Wilson et al. 2021 |
157 | RC-Lot corn-T alex-T michel | 25.1 | 66.8 | Chic-PL-WC | 24.0 | 65.4 | 1.2 |




































