Open Access
Research Article
Max Screen
Copyright: © 2023 Authors Su Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Penicillin and streptomycin are the most commonly used to prevent bacterial infection or contamination in cell culture. However, the role of Penicillin-streptomycin on inducing mouse embryonic stem cells(mESCs) to differentiate into cardiomyocytes is unclear. To illustrate the mechanism of penicillin-streptomycin on inducing mESCs differentiation into cardiomyocytes, we firstly investigated mESCs differentiation into cardiomyocytes by the traditional method. The timespace expressions of cardiac-specific genes were detected by real-time PCR and western blotting at 8 different time-points over a period of 36 days. Furthermore, action potentials were measured by patch clamp for mESC-derived cardiomyocytes. Embryonic bodies through hanging drop method, were incubated with penicillin-streptomycin (P/S) at different concentrations (1×PBS, 1×, 2×, 5×), and then, the number of cardiomyocytes and expression levels of myocardial marker proteins were detected by flow cytometry, cellular immunofluorescence, and western blotting, respectively. We found that the SSEA-1 of mESC presented green fluorescence, and H.E. staining showed karyoplasmic ratio >>1. Besides, TNNI3, TNNT2, ACTN2 and MYL-2v were clearly visible by cellular immunofluorescence. Furthermore, the ACTN2 positive cells rate of 2× and 5× groups were remarkably higher than that in Cntl (1×PBS) group and 1× group (10 Units/mL penicillin; 10 µg/mL streptomycin), as well as the expressions of GATA4, MYL-2v, TNNI3, ACTN2 (p< 0.05). Lastly, the highconcentration penicillin-streptomycin may promote the differentiation of mESC into cardiomyocytes by inducing suppression of the signal pathway for p38/JNK-mediated apoptosis. Our study indicated that P/S promoted mESC differentiating into beating cardiomyocytes and induced up-regulating expression of cardiac-specific markers, suggesting a modulation of hP/S as a potential therapeutic approach for ischemic cardiac disease.
Keywords: mouse embryonic stem cell; Penicillin-Streptomycin; Cardiomyocyte; Differentiation
mESC: mouse embryonic stem cells;
PCR: polymerase chain reaction;
P/S: penicillin-streptomycin;
FCM: flow cytometry;
SSEA-1: stage-specific embryonic antigen-1;
TNNI3: cardiac troponin I;
TNNT2: cardiac troponin T;
ACTN2: α-actinin;
MYL-2v: Myosin light chain-2v;
AP: action potential;
MHC6: Myosin heavy chain 6;
Gja1: Gap junction alpha-1;
Penicillin-streptomycin is composed of penicillin and streptomycin, which are traditional antigram positive coccus and antigram negative bacillus respectively [1-3]. In clinical practice and cell experiments, penicillin and streptomycin are the most commonly used to prevent bacterial infection or contamination [4-6].
At present, the incidence of cardiovascular diseases and the mortality rate increase year by year clinically as the aging of social population and the improvement of living standard [7]. Conventional therapies for heart disease include drugs, interventions, thrombolysis, etc., all are aimed to restore the blood flow to the ischemic zone, but it is still difficult to reverse the patient's cardiac function. Heart transplantation can treat severe heart failure, but it is also limited because of donor origin, immune rejection and other factors [8-10]. With the development of stem cell technology, cell transplantation has attracted more and more attention in recent years, and has gradually been applied to the treatment of myocardial infarction and other diseases [11,12]. Bone marrow derived mesenchymal stem cell is a major source of adult stem cells and it had been once used in the treatment of ischemic cardiomyopathy [13], however, bone marrow MSC is very low in adult bone marrow, and heart disease occurs in middle-aged and elderly people, so this method cannot meet the needs of cell transplantation. Embryonic stem cells are widely used in cardiovascular regeneration due to the ability to differentiate into pluripotent stem cells derived from various types of tissue cells derived from the 3 germ layers of the body [14,15].
Penicillin-Streptomycin is conventionally used for cell cultivation to prevent bacterial contamination. However, few reports have been reported on the induction of myocardial differentiation. Therefore, our study aims to investigate differentiation of mESCs into cardiomyocytes induced by double resistance in vitro and its mechanism to provide a new idea for clinical cell transplantation.
Mouse ESCs (the stem cell center of the Shanghai Institute of life sciences, Chinese Academy of sciences, Shanghai, China) were cultured on irradiated ICR mouse embryonic fibroblasts (inactivated) (Stem Cell Bank/Stem Cell Core Facility, SIBCB, CAS, China) in completed medium (Cyagen, Guangzhou, China) and then, the media of mESC were changed every day.
For differentiation, EBs were formed using conventional hanging-drop method for 6 days as previously mentioned [16].
mESCs were cultured on glass coverslips during the phase of logarithmic growth and then fixed for 10 min in cold acetone/methanol (1:1, V/V) on ice. The cells were performed according to the recommended protocol for use of the BCIP/NBT Alkaline Phosphatase Color Development Kit (Beyotime Institute of Biotechnology, Haimen, China).
The EBs were harvested on days 0, 3, 6, 9, 12, 18, 26, 36, and then, all samples were stored at -80°C in TRIzol (Invitrogen, Carlsbad, CA, USA). According to traditional chloroform-isopropyl alcohol extract RNA, first-strand cDNA was synthesized with the SuperScript III First-Stand Synthesis SuperMix for qPCR (Applied Biosystms, USA). Using cDNA as the template, real-time PCR assays were performed in triplicate using the ABI Prism 7500 HT sequence detection system (Applied Biosystems). The data were normalized to glyceraldehydes-3-phosphate dehydrogenase (Gapdh), and the gene expression levels normalized to that of day-6 cells. The primers utilized are listed in Table 1
The EBs were harvested on days 0, 3, 6, 9, 12, 18, 26, 36 of mESCs or EBs. All samples were extracted according to the manufacturer’s instructions (Invitrogen, USA). The cell homogenates (30 µg) were separated by 10% SDS-polyacrylamide gel electrophoresis and transferred on to a nitrocellulose membrane (Millipore, Germany). The membranes were blocked by 5% (w/v) nonfat dry milk, and then incubated overnight at 4°C with cardiac antibodies; incubated with secondary HRP-IgG antibodies (thermo, USA) for 1 hour at RT. GAPDH was used as an internal control.
The cardiac-specific protein antibodies used for western blotting were as follows: monoclonal anti-cardiac troponin T antibody (1:1000; Abcam); polyclonal anti-troponin I (1:1000; Abcam); myosin light chain 2 polyclonal antibody (1:500; Proteintech Group, Inc., USA); MYH6 polyclonal antibody (1:500; Proteintech) and anti-ACTN2 (1:1000; Abcam); monoclonal anti-GATA4 antibody (1:500; Abcam); monoclonal anti-GJA1 antibody (1:500; Santa Cruz); monoclonal anti-Nkx2.5 antibody (1:500; Santa Cruz); Phospho-p38 MAPK (Thr180/Tyr182) Rabbit mAb (1:1000, CST); p38 MAPK (D13E1) Rabbit mAb (1:1000, CST); Phosphop44/42 MAPK (Erk1/2) (Thr202/Tyr204) Rabbit mAb (1:1000, CST); p44/42 MAPK (Erk1/2) (L34F12) Mouse mAb (1:1000, CST); Phospho-SAPK/JNK (Thr183/Tyr185) Rabbit mAb (1:1000, CST); SAPK/JNK Antibody (1:1000, CST). Monoclonal antiGAPDH antibody (1:1000; Abcam). The secondary antibodies used were: HRP-conjugated goat anti-rabbit IgG (H+L), and HRPconjugated goat anti-mouse IgG(H+L) (1:5000; Thermo).
Cells or EBs were grown on glass coverslips in the absence of gelatin and fixed for 20 min in cold 4% paraformaldehyde. All cells blocked with 5% normal goat serum, were incubated with the primary antibody overnight at 4°C and incubated with the secondary antibody conjugated with FITC- in the absence of bright light. Last, cells were observed with a confocal scanning laser microscope (Sp8; Leica, Germany). Immunostaining with anti-SSEA-1 (1:100; EMD Millipore Co., Massachusetts, USA) and antiOct-4 (1:100; EMD Millipore Co., Massachusetts, USA) was used to verify the undifferentiated mESCs. To verify cardiac differentiation, the following antibodies were used: monoclonal anti-cardiac troponin T antibody (1:100; Abcam); anti-cardiac troponin I antibody (1:100; Abcam); myosin light chain 2 polyclonal antibody (1:100; Proteintech Group, Inc., USA); anti-ACTN2 (1:200; Abcam). The secondary antibody used in this study was fluorescein (FITC)-conjugated AffiniPure goat anti-rabbit IgG (H+L) (1:400; Proteintech). The cell nuclei were counterstained with 0.1% 4', 6-diamidino-2-phenylindole (DAPI; thermo)
EBs cultured on the 22nd day (dispersed condition) were dispersed by 0.1% collagenase type I and 0.1% collagenase type II (Roche) in ADS buffer by stirring. The dispersed EBs were cultured for 2 days in DMEM a glass dish at 37°C supplemented with 10% FBS. The microscope was equipped with a recording chamber and a noise-free heating plate (Microwarm Plate; Kitazato Supply). A 10 mmol/L volume of HEPES was added to the culture medium to maintain the pH of the perfusate at 7.5-7.6.Standard glass microelectrodes that had a DC resistance of 25-35 M when filled with pipette solution (2 mol/L KCl) were used. The electrodes were positioned using a motor-driven micromanipulator (EMM-3SV; Narishige) under optical control.
Spontaneously beating cells were selected as targets, and the action potentials of the targeted cells recorded. Axon patch 200B (Axon Instruments, Sunnyvale, USA) was used for whole cell patch clamp recordings; data acquisition was controlled by Axon Instruments pClamp10 software via a Digidata 1440 acquisition system. Under current-clamp mode, action potentials (APs) of mESC-derived cardiomyocytes were recorded with whole cell patch-clamp configuration in Tyrode’s solution (in mM): 137 NaCl, 5.4 KCl, 1.8 CaCl2, 1MgCl2, 10 Hepes, 10 Glucose, adjusted to pH = 7.4 with NaOH. The glass pipette was filled with the pipette solution (in mM): 120 KCl, 5 EGTA, 1 MgCl2, 5 Na2ATP, 10 HEPES, adjusted to pH = 7.4 with KOH.
Experiments were performed at RT. Data are presented as mean ± Standard Deviation; between-group differences were compared using one-way Analysis of Variance (ANOVA) followed by Student t test. P-values < 0.05 were considered indicative of a statistically significant difference. A minimum of three independent experiments were performed for each analysis. All calculations were analyzed with SPSS Statistics 20 software.
Mouse ES cells formed nodule-shaped or oval colonies with distinct boundaries on the MEFs on the 2nd day (Figure 1. a-c) and then appeared like nested colonies (Figure 1. d). The proliferative mESCs were digested into single cells and hung on inside cover of 150-mm Petri dish in 10% FBS containing differentiation media. These cells continued to proliferate and to gradually thicken and form EBs with globular-shaped structures (Figure 1. e-f).
We found that the cells gradually grew from the edges of the EBs after 24 h of adherent culture to form growth halos which gradually increased in size (Figure 1. g). Furthermore, the cell clusters in the central part of the EBs began to spontaneously beat 30 - 50 times/min after the 16th day of adherent culture (Figure 1. h).
To further confirm that mESCs still remain pluripotent, the cells were rich in ALP and stained bluish violet. Further, the expression of two specific proteins, i.e., SSEA-1 and Oct-4, was examined by immunofluorescence. A vast majority of mESCs were blue-black, and red H.E.-stained cells were rarely observed; these findings implied a karyoplasmic ratio >> 1 in the undifferentiated mESCs. (Figure 2).
To confirm that the EBs continued to be comprised of undifferentiated mESCs, the space-time expression pattern of Isl-1, a marker of cardiac progenitor, was found a wavy-like gene expression pattern of Isl-1 and expressed at a low level at day 9. Interestingly, the expression of Isl-1 was low-level after day 12. These findings indicated that the numbers of new cardiomyocytes remained low. We speculated that none of these genes were highly expressed in the absence of an inducer at any time during the spontaneous differentiation of mESCs into cardiomyocytes. The initial expression of the cardiac transcriptional factors, namely Nkx2.5 and Gata4, were detected on differentiation day 3 and day 6, respectively. And then, both of two transcriptional factors kept up intermediate level. The expression of the cardiac-specific genes Myl-2v, Myh6, Actn2, Tnnt2, and Gja1 were detected initiated between day 9 and day 12, what’s more, their expression levels were subsequently strongly upregulated to varying degrees. It was unexpected that the expression of Tnni3 was detected at day 0 and progressively increased (Figure 4).
To further characterize the space-time expression patterns of the cardiac-specific markers, we examined the expression characteristics of NKX2.5, GATA4, MYL-2v, myosin heavy chain-6 (MYH6), TNNT2, TNNI3, GJA1, and ACTN2 proteins. The expression of NKX2.5, GATA4, MYL-2v, TNNT2, TNNI3, GJA1, and ACTN2 proteins was detected in the early stage followed by a sharp decrease and a gradually increase, thereafter. Interestingly, proteins, such as MYL-2v, TNNT2, TNNI3, GJA1, ACTN2 and MYH6, were rarely observed before day 9. The remaining proteins were expressed at low levels in the early stage, suddenly increased and then kept up with higher level (Figure 4).
When the mESCs-derived cardiomyocytes exhibited robust beating and contraction on the 22th day of differentiation, we detected the expression and location of the markers for cardiomyocytes including MYL-2v, TNNI3, TNNT2 and ACTN2 (Figure 3).
We recorded the action potentials of mESC-derived cardiomyocytes using glass microelectrodes. 22-day-old beating EBs were selected manually and dispersed into small clumps and single cells. The dispersed EBs were cultured for two days before analysis. The microelectrode was advanced to the intracellular cytoplasm and the voltage of the bulk solution and cytoplasm were measured. Rhythmic beating could be detected in the mESC-derived cardiomyocytes. We measured fewer spontaneously beating ventricular-like (n = 12), atrial-like (n = 5) and nodal-like (n = 6) cells due to their relatively lower proportions in mESC-derived cardiomyocytes and because of the difficulty in finding single cells for pacing (Figure 5)
To detect Penicillin-Streptomycin (P/S) playing a role of inducing mESCs differentiation into cardiomyocytes, we found that the expression of the cardiac-specific genes, including cardiac transcriptional factor, Gata4, was markedly increased in the 2× and 5× hP/S groups on the 22th day of differentiation after EBs were treated 30 min by 1×PBS, 1× (10 Units/mL penicillin; 10 μg/mL streptomycin, Gibco), 2×and 5× P/S on day 6 (Figure 6). In addition, real-time PCR showed that it is an optimal time of treatment with 2×P/S that is 30 min (Figure s1).
To further characterize the expression of the cardiac-specific markers, we examined the expression characteristics of GATA4, MYL-2v, TNNT2, TNNI3, GJA1, and ACTN2 proteins in 1×PBS, 1×, 2×and 5× P/S group, which were low-level in 1×PBS and 1× P/S groups, and overexpressed in 2×and 5× P/S groups (Figure 6).
Additionally, we calculated the numbers of beating EBs at four time-points, such as day 12, day 18, day 26 and day 36, and counted the ACTN2-positive cells with FCM in 1×PBS, 1×, 2×and 5× P/S group, respectively. We observed increased numbers of beating ESs in the 2×and 5× P/S groups (Figure 7). Similar results were observed for the numbers of ACTN2-positive cells (Figure7).
Penicillin-streptomycin bind to their receptors, which activate different intracellular signaling components. Interestingly, extracellular signal-regulated kinases (ERKs) are essential for the responses to external environment stimuli. To explore the conceivable role in P/S inducing cardio myogenesis, we detected the expression level of MAPK signal pathway, including p38 MAPK, p42/44 (ERK1/ 2), and JNK. Interestingly, western blot analysis presented that total protein levels of p38, ERK1/2 and JNK were not significantly changed at different concentrations P/S stimuli in mESCs. However, the levels of phosphorylated p38, ERK1/2 and JNK were markedly decreased in P/S-treated mESCs, respectively, especially in 2x and 5x P/S groups (Figure 8). The p-P38 expressions in 2x and 5x groups were decreased more than 30% and 50%, while p-ERK1/2 expressions were decreased more than 75% and 80%, as well as the p-JNK decreased 80% and 60%, respectively, compared to control group (P< 0.05). This suggested that hP/S suppressed phosphorylation of p38, ERK1/2 and JNK, and might reduce p38/JNK-mediated apoptosis.
Due to ethics, law and immune rejection, it is difficult for human embryonic stem cells to be widely used, so mammalian embryonic stem cells are ideal models for studying early embryogenesis, cell differentiation and gene expression regulation [17-19]. The simulation of myocardial cell development in vitro is of great significance to the study of myocardial development model and clinical cardiac drug screening. In vitro experiments showed that ESC differentiation was differentiated into 4 stages [20]: early (d8~d11), metaphase (d12~d15), end stage (d16~19), final end stage (d20~d31). Our experiment first observed that the differentiated beating cardiomyocytes were observed in 13th day, and usually started beating in sixteenth day.
In cell cultural experiments, penicillin and streptomycin (P/S) are the most commonly used to prevent bacterial infection or contamination. However, whether P/S affect cell culture is unclear, especially in the culture of mESCs. Our results demonstrate that high-concentration P/S promotemESCs differentiating into beating cardiomyocytes and induced up-regulating expression of cardiac-specific markers. We have identified the cultured mESC as totipotent: 1, morphological observation: mESC was nested on MEF, with large nuclei, few cytoplasm, dense cell arrangement and clear edges. 2, stem cell surface marker: ESC has the characteristics of expressing early embryonic cell surface antigen SSEA-1, 3, 4, in which mESC only expresses SSEA-1, but does not express SSEA-3, 4 [21,22]. Immunofluorescence results showed that the cultured mESCs were highly expressed SSEA-1 positive (green). 3, ALP and H.E. staining: although ALP and H.E. staining could not prove the pluripotency of ESC, all cells presented obviously bluish violet and were >>1 according to the nuclear to cytoplasmic ratio, which demonstrated cellular undifferentiated states from the side. All these results showed that the mESCs cultured in this experiment was still in undifferentiated state and maintained the totipotent nature of stem cells.
Additionally, whether can mESCs spontaneously differentiate into cardiomyocytes with the complete structural and functional capability in normal culture medium? In this study, cardiac-specific markers, including MYL-2v, MYH6, GJA1, TNNI3, TNNT2, ACTN2 and the two cardiac transcription factors GATA4 and NKX2.5, expressed at different time points, respectively. Interestingly, five proteins, i.e., MYL-2v, TNNI3, TNNT2, GJA1 and ACTN2 were observed at low-level in the undifferentiated mESCs. During cardiomyocyt development, some cardiac genes may express transiently at early embryonic stage. Spontaneous APs were recorded in spontaneously contracting mESC-derived cardiomyocytes. All these data showed that mESCs could spontaneously differentiate into cardiomyocytes.
At present, DMSO, 5-aza, RA, ICA and vitamin C are the main chemical reagents for inducing embryonic stem cells to differentiate into cardiomyocytes, epimedium glycosides and icariin also have such effects [23-26]. Although the traditional inducers can increase the rate of cardiomyocyte production, especially, DMSO has the strongest effect, reaching 78% of the traditional drug, but there are certain side effects of drugs, including toxicity. In this study, the high-concentration P/S was used as the inducer, and it was found for the first time that it has the function of inducing embryonic stem cells to differentiate into cardiomyocytes. The positive rate of ACTN2 positive cells was about 50% in the 2× and 5× groups in contrast to Cntl group maintaining below 10% level. What surprised us is that there is no significant difference between 1× group and Cntl group. At the molecular level, in high-concentration groups (2x, and 5x), cardiac specific proteins, such as MYL-2v, TNNI3, TNNT2, GJA1, ACTN2 and nuclear transcription factor GATA4 began to express highly at 6-day time point.
During mESCs differentiation into cardiomyocytes, wnt/β-Catenin signaling pathway [27,28], MAPK signaling pathway [29,30], PKCε/cTnT signaling pathway and other signaling pathways play important roles [16]. The family of MAPKs plays a key role in cellular responses to extracellular stimulation [31]. The three main families of MAPKs are the extracellular signal-regulated kinase (ERK)1/2 (p42/44), p38, and JNK [32]. Mammalian ERK families are rapidly and transiently activated in response to stimulation [33]. The JNK and p38 pathways have been reported to be critical in induced signaling of cell migration, whereas ERKs respond to mitogen and growth factors that regulate cell proliferation and differentiation [32]. In this study, phosphorylation levels of P38, ERK and JNK were almost at the same time reduced in 2× and 5× P/S groups compared with Cntl group. Therefore, the MAPK signaling pathway may be involved in the process of penicillin-streptomycin induced differentiation of mouse ES cells into cardiomyocytes.
To sum up, during the process of mESCs differentiation to cardiomyocytes, high concentration of P/S can increase the number of beating embryonic bodies, promote the expression of cardiac-specific markers, and accelerate positive proportional of mESCs differentiation into cardiomyocytes for the concentration of P/S through the possible mechanism of suppressing P38/JNK apoptosis signal pathway.
This article is partly supported by the National Natural Science Foundation of China (No. 81700313) and is supported by the fundamental research funds for Tongji University (No. 22120170143).
All authors have no potential conflicts of interest to declare. All authors have no objections to publish the research.

![]()

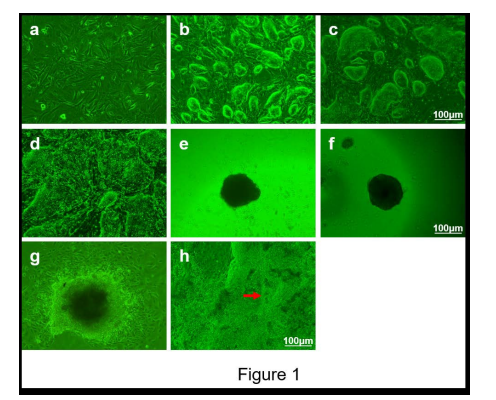 |
| Figure 1: Morphological changes in mouse embryonic fibroblasts (MEFs) and phenotypic characteristics of mESCs. a: The morphology of the MEFs cultured for 24 h. b-d: Colonies of mESCs cultured on MEFs for 2 days, 4 days and 6 days. e-f: Embryoid bodies (EBs) from the hanging drop culture method at 2 days and 4 days. g: Representative EB from the adherent method cultured for 2 days. h: Spontaneously beating cell clusters were found in the cultured cells for 18 days. (the red arrow indicates beating cell clusters). Scale bars=100 μm. |
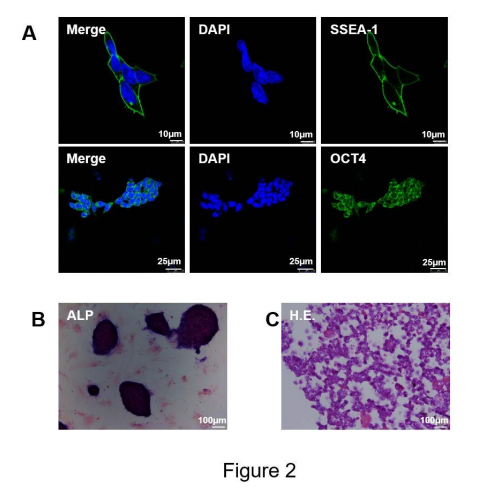 |
| Figure 2: Biochemical characterization of the mESCs. (A): Immunofluorescence staining detecting the expressions of SSEA-1 as well as OCT4, two markers of mESCs. The nucleus was stained by DAPI (blue), and the green showed the positive expression of two markers of mESCs. (B): Alkaline phosphatase staining (ALP) for mESCs showed that the blue-black presented to mESCs. (C): Hematoxylin-eosin staining (H.E.) for mESCs showed that blue nucleus took up almost the entire cell, but red cytoplasm was very rare, which means almost cells were in undifferentiated states. Scale bars: A =10 or 25 μm, B and C=100 μm. |
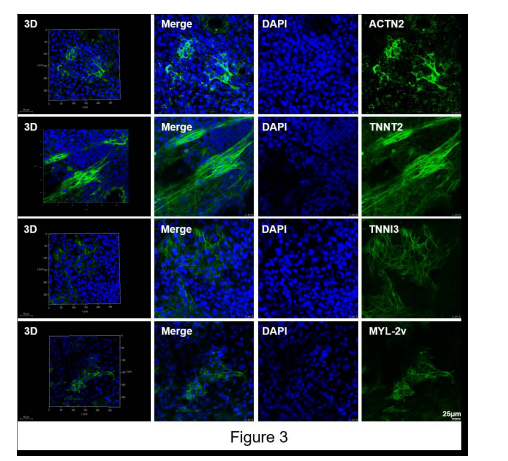 |
| Figure 3: In situ detection of cardiac-specific proteins in differentiated mES cells by immunofluorescence, after 22 days of spontaneous differentiation. Scale bars = 25 μm. |
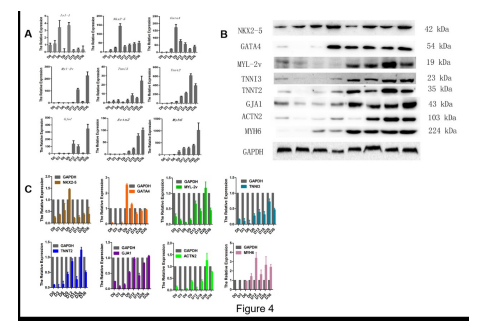 |
| Figure 4: Space-time gene and protein expression analyses performed on the differentiating mESCs. A: Temporal gene expression analyses were performed over 36 days (8 time points) on the cultures of the mESCs differentiating into cardiomyocytes: Isl-1 (cardiac progenitor marker), Nkx2.5 and Gata4 (early cardiac transcription factors), Myl-2v, Mhc6, Gja1, Actn2, Tnni3, Tnnt2 (cardiomyocytes markers). All gene expression levels were normalized to those of the undifferentiated stem cells using the 2-??Ct method and normalized to Gapdh. The day of the first suspension culture was marked as day 0. The samples of undifferentiated and differentiated mESCs were collected synchronously with the samples collected for mRNA. B: The space-time patterns of expression of the cardiac-specific markers are presented (D0-D36). C: The quantitative western blotting data were obtained with Quantity One Software and are summarized as the means ±SD after normalization to GAPDH. |
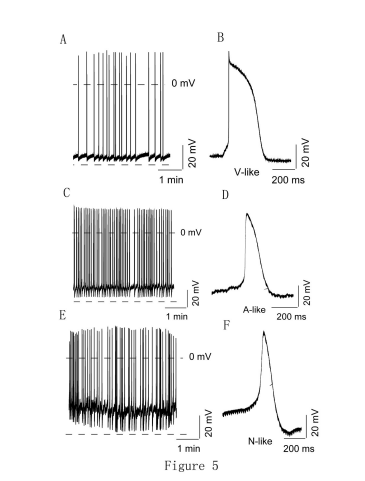 |
| Figure 5: Action potential properties of mESC-derived cardiomyocytes by patch clamp. A-F: Traces of spontaneous action potentials measured in V-like (n=12), N-like (n=6), and A-like (n=5) from cardiomyocytes derived from mESC. |
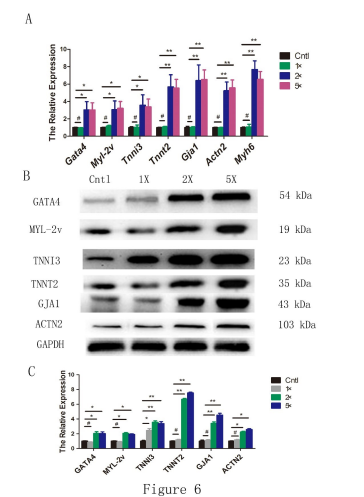 |
| Figure 6: High-concentration Penicillin-Streptomycin (hP/S)promoted cardiomyogenesis, and strengthened the expression of cardiacspecific markers. A: Real-time PCR detection of the effects of P/S on cardiac-specific genes in Cntl (1xPBS), 1x, 2x and 5x P/S group. B-C: Western blot detection of the effects of P/S on cardiac-specific genes at 6-day time point. The proteins were measured as previously described."*": p < O.OS; "**": p < O.Ol; "#": p > O.OS. |
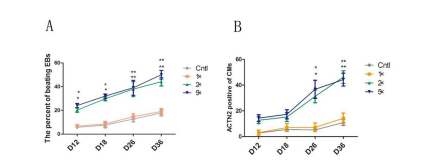 |
| Figure 7: High-concentration Penicillin-Streptomycin (hP/S)promoted cardiomyogenesis, and strengthened the expression of cardiacspecific markers. A: Real-time PCR detection of the effects of P/S on cardiac-specific genes in Cntl (1xPBS), 1x, 2x and 5x P/S group. B-C: Western blot detection of the effects of P/S on cardiac-specific genes at 6-day time point. The proteins were measured as previously described "*": p < O.OS; "**": p < O.Ol; "#": p > O.OS. |
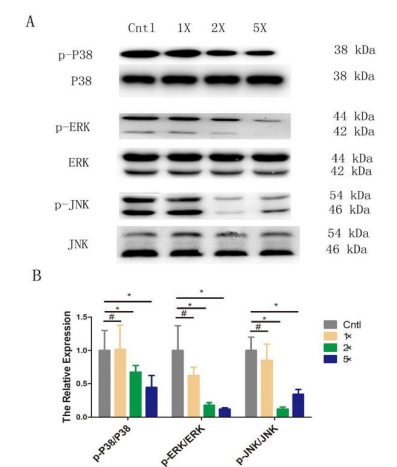 |
| Figure 8: High-concentration Penicillin-Streptomycin (hP/S)promoted cardiomyogenesis, and strengthened the expression of cardiacspecific markers. A: Real-time PCR detection of the effects of P/S on cardiac-specific genes in Cntl (1xPBS), 1x, 2x and 5x P/S group. B-C: Western blot detection of the effects of P/S on cardiac-specific genes at 6-day time point. The proteins were measured as previously described "*": p < O.OS; "**" : p < O.Ol; "#": p > O.OS. |
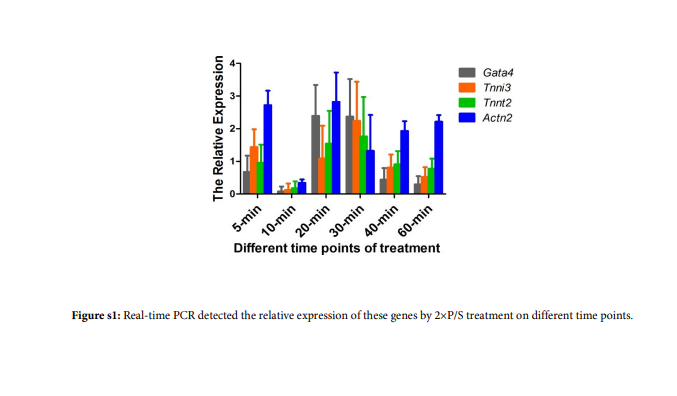 |
| Figure 9: Real-time PCR detected the relative expression of these genes by 2xP/S treatment on different time points. |
Gene name |
Size (bp) |
primers |
Tm ( |
Isl-1 |
109 |
5'-TTTCCCTGTGTGTTGGTTGC-3' |
60 |
|
|
5'-TGATTACACTCCGCACATTTCA-3' |
|
Tnnt2 |
138 |
5'-CAGAGGAGGCCAACGTAGAAG-3' |
60 |
|
|
5'-CTCCATCGGGGATCTTGGGT-3' |
|
Tnni3 |
123 |
5'-AGATATGACGTGGAAGCAAAAGT-3' |
60 |
|
|
5'-TGCAGAGATCCTCACTCTTCG-3' |
|
Mlc-2 |
127 |
5'-CATTCTCAACGCATTCAAGGTG-3' |
60 |
|
|
5'-TGCGAACATCTGGTCGATCTC-3' |
|
Myc6 |
204 |
5'-TGCACTACGGAAACATGAAGTT-3' |
60 |
|
|
5'-CGATGGAATAGTACACTTGCTGT-3' |
|
Gja1 |
111 |
5'-GAAGGATGTGTTAAGCCTCCAA-3' |
60 |
|
|
5'-CTCATTCCCGTAGACAGCAAAG-3' |
|
Gata4 |
88 |
5'-TCAACCGGCCCCTCATTAAG-3' |
60 |
|
|
5'-GTGGTGGTAGTCTGGCAGT-3' |
|
Actn2 |
197 |
5'-CATCGAGGAGGATTTCAGGAAC-3' |
60 |
|
|
5'-CAATCTTGTGGAACCGCATTTT-3' |
|
Gapdh |
213 |
5'-AATGGATTTGGACGCATTGGT-3' |
60 |
|
|
5'-TTTGCACTGGTACGTGTTGAT-3' |
|
Gja5 |
217 |
5'-CCACAGTCATCGGCAAGGTC-3' |
60 |
|
|
5'-CTGAATGGTATCGCACCGGAA-3' |
|
Gjc1 |
198 |
5'-AGATCCACAACCATTCGACATTT-3' |
60 |
|
|
5'-TCCCAGGTACATCACAGAGGG-3' |
|




































