Open Access
Research Article
Max Screen
ISSN: 2639-3336
Copyright: © 2024 Obara Justus Anyieni. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Maize is an economically important crop worldwide. As the world population increases, there is a great need to develop high yielding and stress resistant maize varieties. To provide a future platform for maize improvement, we tested mature embryos as alternative explants for the regeneration of three economically important maize inbred lines: CML331, CML442 and CML444. Callus induction was tested in MS media supplemented with 0, 2, 3 or 4 mg/L 2, 4-D. Maturation of somatic embryos was achieved in hormone free MS medium supplemented with 60 mg/L of sucrose while plantlet regeneration was tested in Naphthaiono acetic Acid (NAA) and 6-Benzyl Amino Purine (BAP) at 0 mg/L of BAP and 0 mg/L of NAA, 0.5 mg/L of BAP and 0 mg/L NAA, 1 mg/L of BAP and 1.2 mg/L of NAA or 1.5 mg/L of BAP and 1.4 mg/L of NAA. CML444 had the highest percentage of callus induction, followed by CML442, while CML331 formed the lowest number of calli. A 2, 4-D concentration of 2 mg/L was optimal for callus induction across the three lines. Our studies revealed that the concentration of BAP and NAA did not impact shoot formation cross the two tested genotypes. CML442 and CML444 successfully regenerated plantlets, while CML331 failed to regenerate. In this work, we demonstrate that mature embryos can be used as alternative explants in in-vitro maize regeneration. The protocol described here was successful and therefore is recommended.
Keywords: Maize Mature Embryos; Callus Somatic Embryogenesis; Regeneration
Approximately 40 million Ha of land in the sub–Saharan region is covered by maize. The consumption rate for more than half of the countries in the region is above 100 g of maize per day per person [1]. Thus, food security in sub–Saharan Africa depends almost solely on this staple crop [2]. The development of high yielding, biotic and abiotic resistant maize varieties is crucial in sustaining production to keep pace with the ever-expanding global population [3]. In-vitro culture and regeneration are unattainable in several maize elite varieties due to inherent genetic differences, rendering them inaccessible to indirect genetic manipulation [4,5]. The genetic make-up of plants is determined by the availability of the stem cells residing in the centre of the meristem that are responsible for new cell generation and initiation of organ formation [6]. Various genes are involved in plant cell division, somatic embryogenesis, and plant regeneration. Inadequate expression of these genes can affect callus regeneration [7]. Explants used in in-vitro maize regeneration have been found to possess a varied responsive ability during laboratory regeneration, which is also connected with their genetic composition, the availability of endogenous hormones and/or their ability to respond to exogenous hormones [8].
Plant regeneration can be achieved via two pathways: direct organogenesis and somatic embryogenesis [9]. Somatic embryogenesis is central to clonal propagation where genetic purity is of great interest [10]. It involves the inoculation of the explant in culture media with subsequent callus induction, callus multiplication, and development of callus into somatic embryos, followed by shoot and root development [11]. Bipolar structures that arise particularly in direct somatic embryogenesis have significant benefits as they have the potential to germinate and regenerate in a single step eliminating the need for serial subculture that might expose the cultures to unwanted mutations [12]. The tremendous connection of root and shoot organs, mainly when plants regenerate from a single somatic cell, is known to preserve genetic integrity as compared to organogenesis [10]. Contrary to other regeneration approaches, it is possible to curb explant growth response to external stimuli through somatic embryogenesis. This enables the explant to only respond to the in-vitro environment conditions that act to reprogram the explant genetic makeup leading to rapid cell differentiation [13]. Further, it is possible to automate and scale up production with somatic embryogenesis particularly, with the deployment of bioreactors, a challenge frequently experienced in clonal propagation [14].
Several explants have been used successfully for maize regeneration including leaves, immature embryos, immature inflorescence, anthers, seedling segments, immature tassels, ovaries, mature embryos, split seeds and shoot tips [15]. Germinated mature embryo-derived explants such as coleoptilar nodes, shoot tips, apical buds, axillary buds and shoot apical meristem have also been used for in-vitro maize regeneration [8, 16]. Despite the availability of various explants, the explant of choice has mostly been the immature embryo [17]. Although they have been intensively used in maize tissue culture, immature embryos remain challenging as embryo extraction is a laborious and time-consuming process. Immature embryos also require a long time to reach the ideal stage for culturing. The need for high precision during pollination and harvesting further complicates their use. Moreover, successful production of immature embryos during winter or cold seasons is difficult [8]. Mature embryo explants, on the other hand, are freely available in large quantities all year round. Mature embryos have been used for regeneration, for instance, [18] successfully regenerated CML216 and Katumani through the mature embryo pathway, an experiment that laid the foundation for this work. Therefore, our primary goal was to evaluate the in-vitro regeneration potential of CML444, CML443 and CML331 mature embryos across different hormonal regimes [17]. These lines were selected because they have a better general and specific combining abilities thus, routinely used in maize breeding. However, the lines are highly susceptible to maize lethal necrosis (MLN) despite them having a central role in breeding purposes. This protocol successfully regenerated two of the tested three lines allowing us to further our research downstream on transformation for MLN resistance or tolerance.
Maize seed material was acquired from CIMMYT-Kiboko, Kenya. Three maize lines were used for regeneration: CML442, CML444 and CML331. Seed multiplication was carried out in Egerton University field three experimental sites. Laboratory experiments were conducted at Egerton University-China Joint Laboratory for Crop Molecular Biology between June, 2020 and March, 2021.
Callus induction was performed on MS medium with vitamins. Media preparation was based on [19] with modifications. Callus induction media was prepared by amending MS media with100 mg/L casein hydrolysate, 2 mg/L L-Proline, 10 mg/L silver nitrate, and 30 g/L sucrose. Various levels of 2, 4-D hormone (0, 2, 3 or 4 mg/L) were incorporated. Media was adjusted to pH5.8 followed by the addition of 3 g/L gelrite. Media was autoclaved at 121˚C for 15 min. 30 ml of the cooling media was dispensed into 100 × 15 mm petri plates under sterile conditions in the laminar flow chamber. Media for callus maintenance was prepared by amending MS media with 2 mg/L 2, 4-D, and 30 g/L sucrose. pH was adjusted to 5.8 followed by the addition of3 g/L gelrite. Media was autoclaved at 121˚C for 15 min. 30 ml of the cooling media was dispensed into 100 × 15 mm petri plates under sterile conditions in the laminar flow chamber. Maturation media was prepared by amending hormone free MS media with 60 g/L and pH adjusted to 5.8. Media was autoclaved at 121˚C for 15 min. 30 ml of the cooling media was dispensed into 100 × 15 mm petri plates under sterile conditions in the laminar flow chamber. The petri plates containing the explants were incubated at 26 ± 2 ˚C under 16 h light and 8 h dark photoperiod and lighting provided by white fluorescent lamps emitting approximately 130 μmol/m2/s of photosynthetically active radiation (PAR) at the surface of the bottles.
MS media was amended with either 0 mg/L of BAP and 0 mg/L of NAA, 0.5 mg/L of BAP and 0 mg/L NAA, 1 mg/L of BAP and 1.2 mg/L of NAA or 1.5 mg/L of BAP and 1.4 mg/L of NAA. 30% g/L of sucrose was added to the different hormone regimes and pH adjusted to 5.8 followed by the addition of 3 g/L gelrite. Media was autoclaved at 121˚C for 15 min. Exactly 40 ml of the cooling media was dispensed into each culture bottle of 100 × 50 mm. The bottles containing the explants were incubated at 26 ± 2 ˚C under 16 h light and 8 h dark photoperiod and lighting provided by white fluorescent lamps emitting approximately 130 μmol/m2/s of photosynthetically active radiation (PAR) at the surface of the bottles
Seeds were surface sterilized following the protocols described [20] and [21] with modifications. Two hundred healthy mature seeds were selected from each maize line. The seeds were soaked for six hours in 50% bleach solution with three drops of 0.01% Tween20. They were then rinsed three times (three minutes for each rinse) using cold distilled water, soaked in 70% ethanol for ten minutes, and then rinsed three times (three minutes for each rinse) with cold autoclaved water. To soften the seeds for easy embryo excision; they were soaked in 40 ml autoclaved called ddH2O in falcon tubes tightly closed with screw caps and wrapped with parafilm. The tubes were then incubated at room temperature for three days.
Using a sterile scalpel, mature embryos were extracted from the maize seeds under a dissecting microscope under the laminar flow chamber. To investigate callus induction in different 2, 4-D hormone concentrations, 20 embryos of approximately 4 mm were placed in petri plates containing MS medium amended with 0, 2, 3 or 4 mg/L of 2, 4-D and incubated in the dark at 26 ± 2°C for two weeks. The experiment was laid in a completely randomized design with three replicates. Two days post inoculation, growing radicle and plumule were excised. After two weeks of callus induction, calli were sub-cultured for four weeks on callus maintenance media. After proliferation, the calli were transferred to petri plates containing maturation medium. Ten calli of about six mm were placed in every plate and incubated at 26 ± 2°C in the dark. Calli were sub-cultured in the maturation medium every two weeks for four weeks. At the end of the maturation period, healthy callus were transferred to the regeneration media.
To test for regenerability, calli were transferred to culture bottles (100 × 50 mm) containing 40 mL of MS media with either 0.5mg/L BAP, 1mg/L BAP and 1.2 mg/L NAA or 1.5 mg/L BAP and 1.4 mg/L NAA. They were incubated at 26 ± 2°C under 16 h light/8 h dark photoperiod with white fluorescent lamps emitting approximately 130 μmol/m2/s of photosynthetically active radiation (PAR) at the surface of the culture bottles. The experiment was laid out in a completely randomized design with three replications.
Individual plants were transferred into three by three-inch pots containing autoclaved, sterile, pre-wetted peat moss. The pots were covered with a plastic dome to contain humidity and placed in a tray (30 cm x 50 cm) with drain holes at 25°C during the day and 20°C at night for three days. Sufficient light illumination was maintained prior to moving the plants to the greenhouse. Domes were then removed, and the plants were transferred to the greenhouse with average temperatures of 32°C during the day and 20°C at night for 4 weeks. The plants were watered once a day and remained in the pots for two weeks. Plants were then transferred to larger pots of about 10×10×20 cm containing a mixture of loam soil with sand and were watered once a day in the greenhouse for two weeks. Our focus was only on callus induction and regeneration therefore; greenhouse data was excluded.
Data on the number of callus formed per line in different 2, 4-D regimes, number of shoots formed per line in different BAP and NAA concentrations, and the number of plants that were able to root was also recorded. Data were subject to general linear model (GLM) using SAS software version 8.2 for analysis of variance. Analysis on significant differences and mean separation of the treatments was conducted using Fisher’s Least Significant Difference test at P≤0.05.
The percentage of callus induction 14 days post inoculation was compared between the lines, and between the different concentrations of 2, 4-D. Callus induction was significantly different between the maize lines. CML444 showed the highest percentage of induction, followed by CML442. Fifty-one percent and 42.5% of CML 444 and CML 442 embryos, respectively, formed calli after 14 days of inoculation. On the other hand, callus induction was significantly low in line CML331. Only 0.75% of CML331 embryos formed calli 14 days after inoculation. (Table 1). Addition of 2 mg/L of 2, 4 D resulted in the highest percentage of callus induction. Increasing the concentration of 2, 4-D to 4 mg/L led to a decrease in the percentage of callus induction (Table 2). The concentration of 2, 4-D also affected callus induction (Table 3). The effect of 2, 4-D concentrations was consistent between the inbred lines CML442 and CML444. In these lines, 2 mg/L of 2, 4-D resulted in the appearance of an average of 9 and 6.33± 0.33 calli three days after inoculation of the embryos. After 14 days of inoculation, an average of 97%±1.33 and 80%±5.77 of inoculated embryos produced calli in CML 444 and CML442, respectively, when the inoculation medium was amended with 2mg/L of 2, 4-D. Callus induction in line CM331 was not affected by the concentration of 2, 4 D (Table 3; Figure 1).
After six weeks of induction and maintenance, calli were transferred to maturation media. Greening of somatic embryos and calli was observed from the 6th day of callus maturation. Some calli in CML442 and CML444 did not form plants instead, produced adventitious roots (Figure 2B). There was also callus greening in both CML444 and CML442 (Figure 2A) however, there was no callus greening in CML331.
A total of 120 CML444, 150 CML442 and 20 CML331 calli were transferred to MS media supplemented with different concentrations of NAA and BAP. Line CML444 had the highest mean of shooting induction followed by CML442 (Figure 3). Shoot induction was not affected by the concentrations of BAP and NAA in either of the maize lines (Figure 4 and Figure 5). Fully developed plantlets were observed three weeks after shoot induction in lines CML442 and CML444 (Figure 6B). In total, seven CML444 and five CML442 calli regenerated plantlets while none of the calli from CML331 formed a plantlet. Plantlets were observed from week 12 (Figure 6A), soma clonal variation or morphological abnormalities were also observed (Figure 6B).
Despite the many challenges, maize regeneration has been predominantly achieved using immature embryos. The ability to form callus in CML442, CML444 and CML331 inbred lines relied on 2, 4-D hormone. Upon callus induction, the 2, 4 D hormone decreases cell wall pressure rendering plant cells water permeable, which causes and increases intracellular water influx leading to doubling of the volume of the embryos size and callus formation. Callus formation would also be influenced by the ability of the explants to express auxin translocation proteins and auxin hormones docking receptors. For instance, the expression of the br2 gene which is responsible for encoding the P-glycoproteins, a hormone central in embryo differentiation, and auxin translocation [22, 23].
Our results showed that there was a higher callus induction rate in lines CML444 and CML442 as compared to line CML331.The ability to form callus would have been due to the presence of auxin in the media [18,11]. In our work, the presence and concentration of 2, 4-D was found to influence callus induction. For instance, CML442 and CML441, had the highest percentage of callus induction in 2 mg/L of 2, 4-D, which was also witnessed in [24]. Increasing the concentration to 3 mg/L resulted in a slight decrease in callus induction in both lines as also observed in [25]. while there was more than 70% decrease in callus induction at 4 mg/L. CML331 formed significantly fewer calli (20 calli) than CML444 (120 calli) and CML442 (150 calli) contrary to [21], where CML331 completely failed to form calli from immature embryo explants
The variation in callus formation between maize lines can be attributed to various factors, including the presence of various endogenous plants growth regulators, the expression of which is thought to be influenced" by in-vitro physical or chemical stimuli [26] and the inherent genetics of the plants. In Arabidopsis thaliana, for example, eight genes are linked to callus formation and are co-expressed with genes involved in plant cell division and cell cycle. The genes are dispersed on chromosomes III, IV, XI, VIII, IX, XII, XV and XVII. Eight quantitative trait loci (QTL) and three epistatic interactions were also found to control type I callus formation in a maize recombinant RIL inbred line [26]. Furthermore, callus formation can be influenced by cellular reprogramming, which is caused by a series of regulatory events that encompass cytokinin, auxin and wound-induced pathways [26].
Appearance of somatic embryos on calli depends on several factors, including the culture environment, and the age of the explant used, with younger explants being the most suitable due to their meristematic properties [23,27,28]. Formation of somatic embryos is also influenced by the ability of the receptor to conform to the target hormone, a scenario that promotes lock and key binding, leading to the docking of the endogenous auxin. Receptor-hormone binding results in rapid cell divisions, which leads to callus formation with subsequent development of somatic embryos [29].
In our study, adventitious roots were observed in the absence of root inducing plant growth regulators, similar to what was reported by [30]. This might have been caused by the ABCG4 gene expression, which is homologous to ABCB14, a hormone that aids in auxin transportation into the roots through the creation of an auxin sink in the root cap. Upregulation of the Eucgr.E01615 (putative expansin-B2) was also found to cause root hair and adventitious root formation during maturation [31].
Elevated sucrose levels and the absence of plant growth regulators during callus maturation dysregulate a number of histone and ribosomal genes that code for hydrolytic enzymes such as nucleases, glucosidases and proteases. Furthermore, it also fails to regulate the α-zein and caleosin storage genes. The two events enhance rapid development of somatic embryos [31]. Moreover, several genes including the ABCG14 gene involved in root and shoot development are upregulated which influence the root to shoot cytokinin translocation via the xylem sap, an event that promotes plant growth [32]. Greening of the CML442 and CML441 somatic embryos causes photosynthesis, hence sufficient nourishment to allow for optimum callus differentiation into plantlet structures. Callus defacement and browning as was observed in CML331 hinder callus development [33] explaining why only green calli possess the regeneration ability.
Regeneration in maize can be induced by the presence and different rates of auxins/ cytokinin hormones [34]. However, our results show that different concentration of exogenous BAP/NAA hormones had no effect on maize regeneration as also witnessed by [18, 13]. This was in agreement with [23] that demonstrated that the presence of endogenous hormones and specific gene expression and suppression patterns are potentially capable of causing plant regeneration in the absence of exogenous phytohormones [35].
Our data also showed that the regeneration frequency of mature embryos was not significantly different from that observed using immature embryos [13]. Genotypic differences are deemed to be key factors that influence the regeneration potential of both immature and mature embryos [36]. Shoot formation could also be controlled by the genetic background that determines the fate of Type II callus formation [11]. In this study, a greater number of regenerants were obtained from embryogenic calli in comparison to organogenic calli, which was contrasting with [21].
Our study also showed that soma clonal variants is common during regeneration via the callus pathway. The event could be caused by the expression of Wus2 genes, which are responsible for inducing active cell division in tissue culture explants and in recalcitrant genotypes. However, overexpression of these genes causes morphological abnormalities also referred to as soma clonal variants [37].
Callus induction was triggered by the presence and different concentrations of exogenous 2, 4-D hormone with intense callus formation activities across the three tested inbred lines being witnessed at 2 mg/L. BAP, or NAA or in combination had no significant effect on shoot formation as plantlet formation also occurred in hormone free media. However, regeneration was significantly different across the two lines probably due to genotypic differences. This study demonstrated that in-vitro regeneration via mature embryo is possible, eliminating the many obstacles experienced with the use of immature embryos. This protocol was successful and could be used for future genetic transformation events.
The authors reported no potential conflict of interest.
Center of Excellence for Sustainable Agriculture & Agribusiness Management (CESAAM), Egerton University.
OJA conceived the study and executed experiment. OJA, KEM, and HGR analyzed the data and wrote the manuscript. JM and OSA contributed to setting up experiments, data analysis, and to develop a structure for the manuscript. All authors have read and agreed to the published version of the manuscript.
The data that support the findings of this study are openly available in figshare at http://doi.org/10.6084/m9.figshare.22707826
We thank Field 3 and the Kenya-China Joint Laboratory for Crop Molecular Biology management for granting us the opportunity to conduct the field and laboratory experiments.

![]()

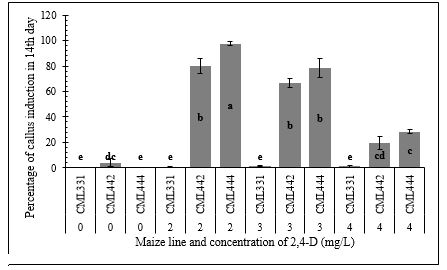 |
| Figure 1: Percentage of callus induction in different concentrations of 2, 4-D hormone in CML331, CML442 and CML444 14 days post embryo inoculation |
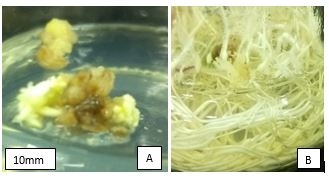 |
| Figure 2: A) Geen regions potential of forming plantlets during callus greening at shoot induction in CML444. B) Rooting observed two weeks after shoot induction in CML444. |
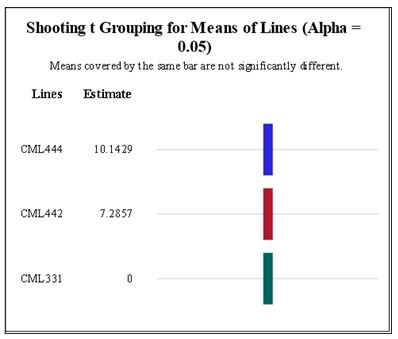 |
| Figure 3: Means of shooting induction in CML 331, CML 442 and CML 444 in all concentrations and respective controls. Lsd0.05=0.581 Means covered with the same bar are not significantly different at p=0.05 |
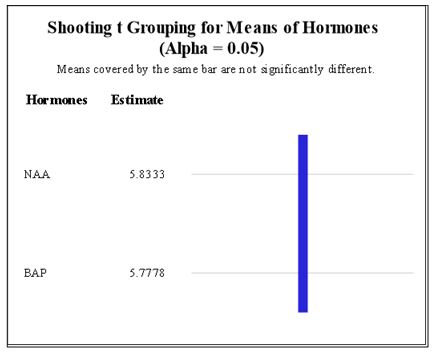 |
| Figure 4: Means of shooting induction at different rates of NAA/BAP for maize lines CML 331, CML 444 and CML 442. Lsd0.05=0.479. Means with same color are not significantly different at p=0.05 |
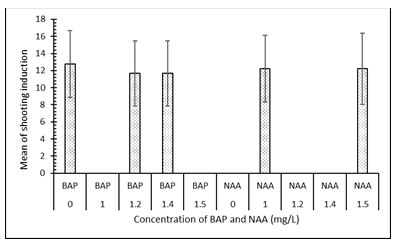 |
| Figure 5: Effect of different concentrations of BAP and NAA on shooting induction in lines CML331, CML442 and CML444 |
 |
| Figure 6: Plantlet regeneration in line CML444. A) A developing plantlet regenerated from mature embryo of line CML444 showing a well-established shoot 14 weeks post callus induction. B) Plantlet regenerated from mature embryo of CML444 showing morphological abnormalities |
Maize line |
Percentage of callus induction |
CML444 |
51.08a |
CML442 |
42.50b |
CML331 |
0.75c |
LSD0.05 |
4.42 |
Concentration of 2,4-D (mg/L) |
Percentage of induction |
0 |
1.33d |
2 |
59.44a |
3 |
48.78b |
4 |
16.22c |
LSD0.05 |
5.11 |
Concentration of 2,4-D (mg/L) |
Maize line |
Percentage callus induction |
0 |
CML331 |
0 ± 0.00 |
0 |
CML442 |
4 ± 3.06 |
0 |
CML444 |
0 ± 0.00 |
2 |
CML331 |
0.67 ± 0.67 |
2 |
CML442 |
80 ± 5.77 |
2 |
CML444 |
97.67 ± 1.33 |
3 |
CML331 |
1.33 ± 0.33 |
3 |
CML442 |
66.67 ± 3.33 |
3 |
CML444 |
78.33 ± 7.36 |
4 |
CML331 |
1 ± 0.58 |
4 |
CML442 |
19.33 ± 5.21 |
4 |
CML444 |
28.33 ± 1.67 |




































