Open Access
Research Article
Max Screen
ISSN: 2639-3336
Copyright: © 2023 Jane Roche. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Stress of the endoplasmic reticulum (ER) is induced by the accumulation of misfolded proteins in this organelle. It can be triggered by biotic and abiotic stress (particularly heat), but also by chemical treatments (such as DTT and tunicamycin) at a laboratory scale. To respond to this stress, various cellular mechanisms are involved, including the highly conserved protein repair pathway, the UPR (Unfolded Protein Response). This pathway aims to restore protein homeostasis in cells. In cereals, seed storage proteins (SSP) represent one of the characteristics that determine grain quality and are of great interest to agriculture. Unlike dicots, the dynamics of the UPR induction pathway in monocots are poorly documented in the literature.
In this review, in addition to the different mechanisms involved in the protein homeostasis maintenance in response to ERstress in cereals, orthologs of UPR induction’s markers have been identified. A focus has been made on the dynamics of the UPR pathway in seedlings and in seeds under chemical UPR inducers (DTT and tunicamycine) and heat stress. The interactions between the different actors of the UPR pathway are also reviewed and discussed.
Keywords: UPR; Cereals; Environmental Stresses; Dynamics
ER: Endoplasmic reticulum; ERQC: ER Quality Control; ERAD: ER-Associated Degradation; IRE: Inositol-Requiring Enzyme1; RIDD: Regulated-IRE1 dependent decay; UPR: Unfolded Protein Response; DTT: Dithiothreitol; TM: tunicamycin; PERK: Protein Kinase RNA-Like Endoplasmic Reticulum Kinase; bZIP: basic leucine zipper transcription factor; ATF6: Activating transcription factor 6; TMD: Transmembrane domain; S1P: Site 1 Protease; BiP: Binding Protein; COP2: Coat Protein Complex II; S2P: Site 2 Protease; ERSE: ER-stress response element; pUPRE: plant UPR element; XBP1: X-box binding protein 1; NLS: nuclear localization signal; bZIP60u: unspliced form of bZIP60; bZIP60s: spliced form of bZIP60; mUPRE: - mammal UPR element; PDI: Protein Disulfide Isomerase; ERDj: Endoplasmic reticulum (ER)-resident J-domain-containing proteins; PB: Protein Bodies; Hrd: HMG-CoA reductase degradation protein; CNX: Calnexin; CRT: Calreticulin; NAC: NAM-ATAF-CUC2 transcription factor; NTL: NAC with transmembrane motif 1 – Like; PDIl: Protein Disulfide Isomerase like; ERO1: ER Oxidoreductin 1; DAA: Day After Anthesis; SSP: Seed Storage Protein; DAF: Day After Flowering; LT: Low Temperature; HT: High Temperature; MLG: Mix-linkage Glucan
Cereals have a major economic value in the world, with more than 2764 million tons produced worldwide (mainly corn, wheat, and rice) (Fao.org). They are an important part of the human diet, but are also used to produce animal feed and biofuels. An increase in global temperature between 1.5 and 5 °C is predicted to occur in 2100 (IPCC Report 2021). Cereals are temperature-sensitive plants, mainly during their reproductive phase [1,2]. In general, heat stress leads to a decrease in photosynthetic activity correlated with an increase in leaf senescence due to a decrease in photosystem II activity, chlorophyll content, and stomatal conductance [3]. These perturbations lead to a decrease in yield components, such as the number of grains per spike [4]. The grain quality (starch content for example) is also affected; due to the reduction of enzymes involved in MLG (Mix-linkage Glucan) synthesis such as cellulose synthase like-F6, which are heat sensitive [5-8]. Furthermore, the duration of the cellularization and grain filling stage also decreases, leading to a lower grain weight and size (thickness, length, width) [8,9].
At the cellular level, heat stress induces, among other response phenomena, the disruption of protein homeostasis in the endoplasmic reticulum (ER) mainly due to an increase in the load of unfolded proteins, which constitutes a stress called ER-stress [10]. To limit this stress, restore homeostasis and avoid apoptosis, plants trigger specific molecular processes [11]. Indeed, in the ER organelle, unfolded proteins are identified by a protein quality control mechanism called ER Quality Control (ERQC) and are degraded by ERAD (ER-Associated Degradation). The expression of genes that encode proteins involved in these two mechanisms increases in response to ER-stress [12,13]. In the primary response to ER-stress, in cereals, as in dicots, RNAse activity of an enzyme called IRE1 (Inositol Requiring Enzyme1) is induced, leading to the degradation of mRNAs to limit neosynthetized proteins. This mechanism is called Regulated-IRE1 dependent decay (RIDD) [11,14,15]. Some authors suggest that some mRNAs degraded by the RIDD mechanism would be anti-autophagy mRNAs and their degradation would activate the pro-survival autophagy pathway, which, in dicots, eliminates a part of the ER containing misfolded proteins in autophagic bodies [11,16,17]. In parallel to this pro-survival pathway, the UPR (Unfolded Protein Response) pathway is activated to repair misfolded proteins by decreasing folding demand, upregulating protein folding capacity, and ability to degrade proteins [5,11,18,19].
The UPR is an ER-specific signaling pathway that has been extensively studied in many living organisms, notably through the use of chemical inducers such as DTT and tunicamycin (TM). These molecules are considered as UPR inducers because they are responsible, respectively, for inhibition of disulfide bridge formation [20] and N-linked glycosylation [21], post-translational modifications that are essential for correct protein folding [22].
Furthermore, the UPR pathway has been described to be involved in the response of cereals to abiotic stresses [2,5,19].
The UPR pathway is present in mammals [23], yeast [24], and plants (dicots [25,26]; and monocots [2,19,27]) to reduce ER-stress. In plants, one of the UPR induction arms, corresponding to a mammalian protein called PERK, is lost [20], but two different induction arms involving two transcription factors bZIP (basic leucine zipper) that carry a transmembrane domain, bZIP28 and bZIP60 are conserved [5,19,25,26].
In plants, 1 or 2 orthologs of ATF6 (Activating transcription factor 6) from metazoans have been identified [28-30] such as AtbZIP28 and AtbZIP17 in Arabidopsis thaliana (At3g10800 and At2g40950, respectively [26,31]). Among cereals, there are two transcription factors acting separately, for example in rice: OsbZIP39 (accession number Os05g0411300) and OsbZIP60 (accession number Os07g0644100) [6,32] and in Chenopodium quinoa: CqbZIP67 and CqbZIP44 [33] while sometimes only one ortholog is described (for example, in maize: ZmbZIP91/ZmbZIP17 (accession number Zm00001d007042) [11,34]). When these two genes have distinct sequences, their participation in the response to abiotic stresses can vary depending on the type of stress. For example, in Arabidopsis thaliana, AtbZIP17 expression is induced in response to salt stress [26,35], while AtbZIP28 is specifically induced in plants subjected to heat stress or to DTT and TM [26,31].
The transcription factor bZIP28 (which is most commonly studied) is a transmembrane protein located under normal growth plant conditions in the ER membrane [11,27,34]. In addition to its transmembrane localization domain (TMD), it has a transcriptional activation domain in the cytosolic N-terminal domain of the protein [27,32]. It is expressed in all tissues and constitutively under all experimental conditions tested in the literature [27].
In cereals, in response to ER-stress, the transcript level of this transcription factor (ZmbZIP17 in maize, for example) is amplified [11,27,32,34]. This increase occurs during the first hours of chemical treatment inducing the UPR pathway (2 and 6 h of 2 g/mL TM and 2 mM DTT treatments) and the transcript level decreases afterwards (12 h) [34]. In fact, under ER-stress, the quantity of misfolded proteins increases. To maintain protein homeostasis, a chaperone protein called BiP (Binding Protein) originally associated with bZIP28 binds misfolded proteins, leading to its separation from bZIP28 [36]. This phenomenon results in the detachment of bZIP28 from the ER membrane. It is then integrated into COP2 vesicles to undergo two successive proteolytic cleavages in the Golgi apparatus by proteases at their specific sites S1P and S2P (protease site 1 and site 2) [37]. Following this proteolysis, bZIP28 is redirected into the nucleus where it binds to ERSE1 cis-elements (5'-CCAAT-N10-CACG-3'), specifically to the CACG sequence, and strongly to pUPRE2 (5'- GATGACGCGTAC-3') of ER-stress responsive genes promoters [32,37]. This binding results in the induction of the transcription of genes encoding proteins involved in protein folding machinery, ERQC, protein secretion, and ERAD to decrease the number of misfolded proteins [11,13,27,32,34,38]. The presence of a specific cleavage site of a protease localized in the Golgi apparatus (called S1P) [13,27,32,34] and the fact that in cereal protoplasts, bZIP28 without transiently expressed TMD is located in the nucleus under normal plant growth conditions [27], indicates the same ability to transfer from the ER membrane to the nucleus than in mammals [23] and dicots [36,37]. The study of ZmbZIP17 overexpression lines, showed that this transcription factor is also involved in ER-stress tolerance by regulating seedling morphogenesis and improving the growth [34].
The transcription factor bZIP28 is involved in the negative autoregulation of its transcript to maintain protein homeostasis. This was highlighted by the use of transgenic lines overexpressing OsbZIP39 truncated at the TMD, showing a decrease in the OsbZIP39 form [27].
Numerous orthologs of Arabidopsis thaliana IRE1 (AtIRE1a and AtIRE1b) [39], belonging to the second arm of UPR induction (IRE1/bZIP60), have been identified in cereals (OsIRE1 in rice (accession number: Os07g0471000) [10,14,15,32,40]; ZmIRE1 [11] in maize and BdIRE1 in Brachypodium distachyon [5]).
Similarly, orthologs of bZIP60 in Arabidopsis thaliana (AtbZIP60: At1g4299039) [41], itself an ortholog of mammalian XBP1 [23], have been identified in cereals by sequence alignment (OsbZIP74/50 in rice (accession number: Os06g0622700) [6,10,14,32,38,42-44]; ZmbZIP60 in maize (accession number: Zm00001d046718) [2,11,45]; TabZIP60 in wheat (accession number: TraesCS7A02G398400.1 ;TraesCS7D02G392800.1 and TraesCS7B02G299200.1) [19]; CqbZIP81/92 in Chenopodium quinoa [33]; BdbZIP60 in Brachypodium distachyon (accession number: Bradi1g35790) [5]; SbbZIP60 in sorghum (accession number: XP_002437297) [10,46] or HvbZIP60 in barley (accession number: BAJ96708) [10]).
IRE1 is a transmembrane protein localized in the ER membrane that is structured in 2 monomers in dicot and monocot species [10,40,47-49]. Under ER-stress, the kinase domain of one monomer autotransphosphorylates the other, activating the RNAse domain [10,14,39,40,47,48,50].
In mammals, yeast and dicots, under normal growth conditions, IRE1 protein is bound to the chaperone protein BiP, which serves as a monomer holding the protein in the ER membrane [39,47,50-52]. Upon accumulation of misfolded proteins, BiP detaches from IRE1 inducing dimerization of the IRE1 protein and its activation [10,14,32,39,47-51]. As in other organisms [16,24,49,53,54], in cereals, the RNAse activity, associated with the RIDD activity of IRE1, is also involved in the unconventional splicing of bZIP60 mRNA [11,14,15]. The spliced region does not have the typical characteristics of the spliceosome (GU-AG) [11,32], but it is possible by recognizing the double loop stem structure of its mRNA containing a CxGxxG sequence in each loop [5,10,19,32,45,55]. Unlike dicots, the bridge size in cereals between the two loops is shortened by 3 bases. Thus, the eliminated intron is about 20 bases whereas it is 23 bases in Arabidopsis thaliana [5,10,19,45]. This splicing leads to a shift in the reading frame, creating the spliced form with no longer a C-terminal TMD sequence but an NLS sequence instead (nuclear addressing signal sequence), while maintaining a functional N-terminal part [19,32,45]. The primary role of IRE1 in the splicing of bZIP60 in cereals has been demonstrated by the use of IRE1 KO mutants that do not express the spliced form of bZIP60 under stress conditions; and by the use of overexpression mutants of IRE1 that constitutively express bZIP60 spliced even under normal growth conditions [10,32].
In cereals, as in dicots [25,51], the unspliced form of bZIP60 (bZIP60u) transcript that contains a transmembrane domain is constitutively expressed under normal growth conditions, as well as under ER-stress inducing conditions [5,10,19,32,45]. This domain leads to its localization exclusively in the ER membrane. However, in plants in general, during an ER-stress, an increase in the amount of bZIP60u transcripts is observed in the cytoplasm at the same time as the appearance of the spliced form (bZIP60s) in the nucleus [5,10,19,32,38,43,45]. In rice specifically, once in this organelle, bZIP60s binds directly, and independently of other transcription factors, to the cis-element pUPRE2 (5'-GATGACGCGTAC-3') of promoters of genes coding proteins involved in the control of protein folding [38]. This cis-element differ from the mammalian mUPRE (5'-TGACGTGG-3') with the exception of the TGACG part [56], and from the UPRE motif of the plants identified in Arabidopsis thaliana, pUPRE (5'-ATTGGTCCACGTCATC-3') [57]. A mutation in the pUPRE2 motif greatly reduces the binding capacity of the transcription factor bZIP60s, therefore, reducing the expression of genes related to the promoter containing this cis-element [38]. The binding of bZIP60s induces the transcription of genes encoding proteins involved in the restoration and maintenance of protein homeostasis; such as PDI (disulfide bridge formation, and protein bodies formation), bZIP60 (over-induction of the UPR arm), BiP1 (involved in folding,chaperone protein, and protein bodies formation), ERDj7 (involved in quality control, ERQC and protein body formation), Hrd3 (involved in ERAD), Derlin1 (involved in protein degradation by ERAD), Erdj3 (involved in binding and activation of BIP), CNX (Calnexin constituting the folding complex) and CRT (Calreticulin constituting the folding complex) [5,11,20,32,38,42,43,58-62].
bZIP60s KO mutants in maize (5'UTR insertion of a transposon) and Arabidopsis thaliana mutants complemented by the bread wheat gene (TabZIP60s) indicate that bZIP60s, is involved in the transcription induction of canonical UPR genes, but also plays a role in temperature tolerance [2,19]. When bZIP60s expression decreases, vegetative growth is negatively affected [2] and, when it is overexpressed, a significantly higher rate of germination and survival is observed in response to heat stress compared to wild plants [19].
In addition, the spliced form of bZIP60 is involved in a positive feedback loop, because it is self-regulated in response to ER-stress by binding to its own promoter to induce its transcription [19,42]. In rice, another transcription factor of the NAC family (OsNTL3) has been described to positively regulate bZIP60. Localized in the plasma membrane through a TMD domain, this transcription factor (OsNTL3) is induced during ER-stress and relocated to the nucleus where it binds to promoters of genes involved in protein folding (BiP, Calreticulin, PDI) [42]. Liu et al. have also demonstrated that OsNTL3 KO mutants have a lower survival rate in response to ER-stress induced by a high temperature stress [42]. Furthermore, the expression of OsNTL3 is also positively controlled by OsbZIP74, which amplifies the ER-stress response [42]. Finally, proteolysis targeted by the 26S proteasome and ERAD pathway has been observed to be involved in the balance of the abundance of bZIP60 proteins under normal plant growth conditions and under heat stress to limit the excessive accumulation of these proteins. During heat stress and during recovery from this stress, a strong increase in the level of bZIP60s protein is observed in response to treatment with MG132 (a proteasome 26S inhibitor) [19].
The transcription factors bZIP28 and bZIP60s are induced in response to ER-stress and bind to the particular cis-elements pUPRE2, a motif they share [5,27,38,45]. Their requirements for transcriptional activation linked to this cis-motif vary and may involve other unknown factors [38]. Some authors have shown that bZIP28 induces the expression of the transcription factor bZIP60 [11,27]. Indeed, the UPR arm involving bZIP60 appears to be activated later than bZIP28 [27,32]. So bZIP28 would participate in a faster response than the arm it amplifies, IRE1/bZIP60 [11,27,32]. Thus, Takahashi et al. propose the existence of a regulation phase mediated by bZIP28 alone during short stress and the participation of bZIP60 during prolonged stress to obtain a more robust and lasting response because of the positive self-regulation of bZIP60 [27,32,42].
Furthermore, in response to ER-stress, the transcription of some genes (OsBIP2, 3, 4, 5) is completely inhibited in bZIP60s KO mutants, while others (OsBiP1, CNX, PDIl1-1, Erdj3, ERO1) are only less expressed [5,32,38]. These observations support the hypothesis that, depending on the type of genes induced in response to ER-stress, their expression driven by bZIP60s may be dependent on the presence of bZIP28. Therefore, the expression of some response genes is modulated by the presence or absence of the two transcription factors, while others require only the presence of bZIP60s [5,11,14,27,32,38]. Thus, in monocots, as in dicots [31], these observations suggests that bZIP28 and bZIP60s could regulate UPR pathway genes separately and also cooperatively through the formation of heterodimers on the cis-elements of the promoters of these genes [32,38].
In addition to that, in response to ER-stress, the RIDD mechanism is induced to limit the load of the translation machinery [15], but this induction does not reduce the overall rate of translation initiation, only the overall transcript quantity [20]. In cereals, translation is not selective and is done in proportion to the abundance of mRNA relative to the amount of total RNA [20]. For this reason, the high amount of UPR gene transcripts induced by the two induction arms are translated into higher amounts than other mRNAs. Therefore, to limit the overload of the translation machinery in the already stressed ER, a decrease in translation efficiency (number of translated mRNAs compared to transcribed RNAs) is observed, including the UPR genes [20]. In maize, the mRNAs not supported by the polysomes are stored in abundance in granular stresses. This storage is proportional to their amount in relation to the quantity of total RNA; suggesting the formation of an mRNA reserve that could later enter the translation machinery [11,20].
Considering the complexity of the interactions between the bZIP28 and bZIP60 arms, we chose to consider only the IRE1/bZIP60 arm in the induction of the UPR pathway. The IRE1/bZIP60 arm can be artificially induced in the laboratory using chemical inducers such as DTT and Tunicamycin [45] but also in response to biotic [10,51], and abiotic stress [35,51]. In this paper, we decided to review the variation in the expression of the bZIP60 gene of cereals considered as a master gene of the UPR pathway, and UPR effectors (such as BiP, PDI, ERO1, CNX, CRT, ERDj proteins) in the different parts of the plant in cereals under chemical treatment (DTT and Tm) and under heat stress, as the most important factor reported inducing bZIP60 splicing [19,45]. Like Vitale and Pedrazzini, we paid a special attention to the seed compartment since cereal grains contain between 10 and 15% protein (SSP) that transit through the organelle of the ER, particularly during the grain filling when SSP are synthesized [63,64].
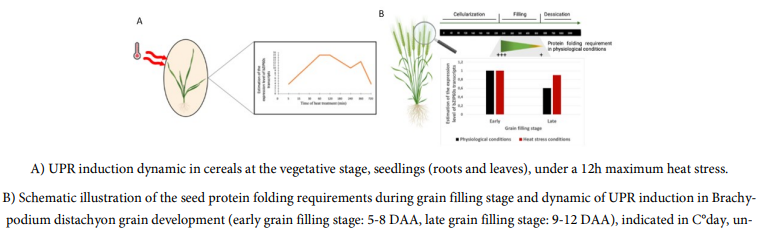
Unlike the unspliced form of bZIP60, which is expressed in any seedling tissue under normal growth conditions, the spliced form of bZIP60 is not detected [5,19,45]. This is consistent with the fact that under physiological conditions, the IRE1 enzyme, inactivated by binding to the chaperone protein BiP, did not initiate the unconventional splicing of bZIP60 [47]. In this review, data from different publications on cereals were summarized in order to be compared (Fig1 and Fig2)
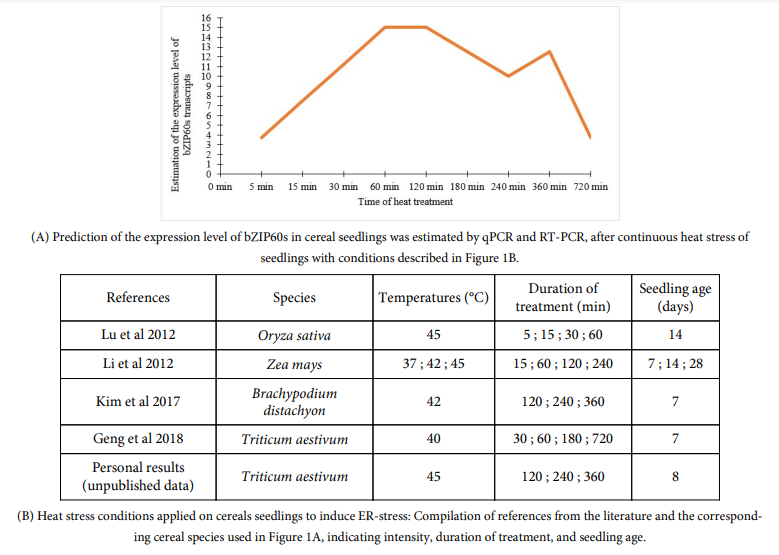
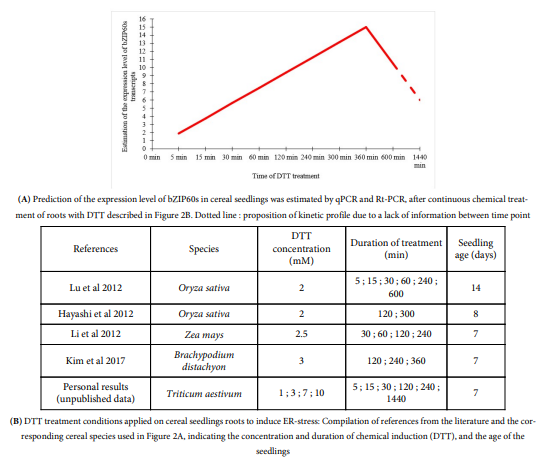
During ER-stress induced by a chemical treatment (DTT or TM) or by heat stress, a bZIP60 accumulation is generally observed [5,10,19,42,45] ([6] DTT: 2 mM; [11] TM: 5 µg/mL). Furthermore, as in dicots [65], the amount of bZIP60s transcripts in cereals changes over treatment time [5,19] and differently over time depending on the treatment applied [5]. For example, in response to heat stress, it is expressed after 5 min of treatment [10] and a first peak is observed around 1-2 h, followed by a slight decrease (Fig 1) [5,10,19,45]. Then, a second lower peak of expression is observed around 6 h before decreasing again (Fig 1) [5,19]. In response to DTT treatment, a rapid induction is observed, also around 5 min [10], but followed by a high increase in the amount of bZIP60s transcripts up to 6 h after treatment (Fig 2) [5,45].
Unpublished personal data obtained in our laboratory suggest a decrease in bZIP60s transcripts after 24 h of DTT treatment (Fig 2). TM treatment induces the UPR pathway less intensively than DTT [5,45]; with a later induction of the spliced form of bZIP60, between 3 and 4 hours [5,10,11,45]. The intensity of the applied treatment also plays a role in this kinetics [45]. In fact, Li et al. showed by semiquantitative PCR that the accumulation of bZIP60s transcripts is proportional to the intensity of heat treatment (37, 42 and 45 °C) [45]. Subsequently, other authors evaluated the expression level of bZIP60s by qRT-PCR by modulating the intensity of heat treatment (31, 33, 35 and 37 °C) and found that its expression was multiplied by 19 from 31 and 37 °C [2].
Similar kinetics of response were identified between vegetative and root tissues [5,19]. The age of the seedlings and tissues appears to also be important in the ER-stress response. Indeed, 7 days old seedlings or young tissues (basal region and transition leaves) have a more intense response to heat stress compared to older tissues or individuals (2 and 4 weeks, maturing and mature leaves) [45]. Finally, after returning to ambient temperature, a rapid decrease in the spliced form of bZIP60 was observed in seedlings, followed by a total disappearance after 2 h [45]; this corresponds to the rapid return observed in dicots in the inactivated state of IRE1 by binding BiP again after stress [47].
In cereals, grain maturation is divided into two stages before dessication: cellularization (3-5 DAA in Brachypodium) and filling stages (5-18 DAA in Brachypodium) [66]. During the grain filling stage, there is a strong synthesis and accumulation of proteins in the grain ER [8,60-63,67,68]. These proteins are: i) metabolic, related to the synthesis of polysaccharides such as enzymes synthases of complex polysaccharides (mainly starch); ii) storage (SSP: Seed Storage Proteins) used as a source of nitrogen and carbon during germination (80-85% of the grain proteins) [5,8,13,60,64,69]. This increase in protein synthesis leads to an overload of the translation machinery [5,7,18,69]. Such as in the other parts of the plant, this phenomenon triggers several cellular processes such as ERQC, ERAD, and UPR, to restore protein homeostasis in the grain and maintain correct filling [5,7,12,13,18,60-62,69,70]. As described in the review of Vitale and Pedrazzini, an overproduction of SSP does not induce the spliced form of bZIP60 at the same intensity depending on the type of SSP [18,63]. For example, Brocca et al. demonstrated different overexpression levels of bZIP60s in Arabidopsis mutants expressing two types of maize storage proteins (yzein 16kDa and 27kDa). The authors found that mutants expressing low molecular weight zein (16kDa) had a higher relative level of expression of bZIP60s (3x) than those expressing a high molecular weight protein. This difference in expression may depend on the affinity of SSPs for the chaperone protein BiP involved in their folding [18,63,70].
Under physiological growth conditions, during wheat seed pre-filling stage (10 DAA), there is an important SSP degradation rate (around 25% of the neosynthetized protein), possibly by the action of ERAD [67]. Experiments performed on rice under an MG132 treatment, some cysteine-rich 13 kDa prolamin polyubiquitinated were observed [12]. At early stage of wheat grain filling (14 DAA) an SSP mRNA accumulation is detected in the grain with a high protein synthesis rate leading to a fast SSP accumulation in the endosperm in development (17 DAA) [67]. In these conditions, to limit excessive stress in the ER, SSP aggregates by intra/inter molecular disulfide and hydrophobic bonds, thanks to PDI and BiP actions, into heteropolymers, called protein bodies (PB) [12,59-62,71-73]. There are 2 types of PB, PB1 (derived from ER), PB2 (transport to the vacuole) characterized by a different composition of SSP [13,59,61,62,68,73,74]. For the same reasons, the ERAD and UPR pathways are also induced during grain filling [5-8,12,43,69,70]. Regulation of the genes involved in these processes (PDI (disulfide bridge formation and PB formation), bZIP60 (UPR pathway induction), BiP1 (folding-involved, chaperone protein and PB formation), ERDj7 (quality control, ERQC involved in PB formation), Hrd3 (ERAD-involved), Derlin1 (ERAD protein degradation), Erdj3 (BIP binding and activation), CNX (folding complex) and CRT (folding complex)) differs depending on the stage of grain development [5,6,7,12,13,69]. Their expression level is higher in the early stages around 5-14 DAF compared to the late stages, around 20 DAF in Brachypodium distachyon [5,6,7,12,13,69]. For example, Shimoni et al. showed that PDI quantity is increased in wheat grain several days (5-9 DAA) before the beginning of SSP accumulation (17 DAA) and decreased during late seed developmental stage (after 21 DAA) [59]. Therefore, the UPR pathway is more strongly induced during the early stages of grain filling under normal plant growth conditions [5,6,69]. The production of polysaccharides in the grain decreases during its development, probably because of the lower quantity of polysaccharide synthases in the late stages of grain filling which may be under of the ER folding control [5,8]. The requirements for the folding of neosynthesized proteins are lower when the endosperm filling ends; therefore, the decrease in UPR induction during grain development would coincide with the protein folding requirements [5,7]. Thus, Kim et al. suggested that UPR would be necessary for the correct maturation of grain proteins such as SSPs or polysaccharide synthases, during endosperm development. Indeed, successful grain filling would be strongly correlated with the cell ability to respond to ER-stress under normal grain development conditions [5,6,7,12,43,63,69,70].
In addition, Wakasa et al. have shown that an optimal accumulation of BiP1 in rice coincides with an increase in proteins involved in folding, which significantly improves the content of storage proteins [6]. Indeed, the perturbation of the expression of proteins involved in the protein folding machinery in the ER (such as CNX, CRT, PDI) in BiP1 overexpressing or KO mutants in rice provoked an abnormal phenotype because of ER-stress (i.e., decrease of yield and quality traits of the grains such as size, length, width, thickness, smaller weight, floury features, and a decrease of seed storage protein production and starch accumulation) [6,7,8,9,43,75].
In cereals, ER-stress in grain has been described following the application of heat stress [5]. The ER-stress induced a decrease in grain weight and may be due to a smaller amount of starch and fewer starch granules with a more fragile structure; or a decrease in the starch synthases [5,6,8,75]. Furthermore, the amount and accumulation of SSP decreased [5,6,15,43,67,75]. Several hypotheses have been proposed to explain this reduction: the RIDD mechanism that would target specific SSPs [15], the ERAD mechanism may degrade a higher amount of misfolded proteins [12,67], the reduction of the grain filling stage [5] and its duration that could be observed in response to the application of moderate temperatures ([9] LT: 21/15 °C, HT: 29/23 °C). In transgenic rice overexpressing or underexpressing BiP1, ER-stress induces an increase in the transcriptional activation of some ERQC, ERAD and UPR genes during seed development with a greater induction in the early stages [6,43] ([70] DTT: 2 mM/TM: 5 µg/mL). During a heat stress of 30 °C for 1 week, the expression profile of bZIP60u and bZIP60s are similar to the controlled condition (22°C), i.e., a decrease in the late stages compared to the early stages [5]. However, an expression of bZIP60u and bZIP60s is observed to be more important during heat stress in the late stage compared to the physiological condition [5]. On the contrary, the level of transcripts of only two genes, BiP and PDI, in the early stages is lower in response to heat stress, and there is a decrease in the late stages compared to the early stages under both conditions. Therefore, the early stages of grain filling may be more vulnerable to heat stress because there is a high accumulation of SSP (leading to ER-stress) during these stages. It has been proposed that prolonged stress inducing a higher ER-stress level, such as in the early stages of grain filling, could induce other actors of the UPR pathway, such as BiP2, 3, 4, 5 [5].
The endoplasmic reticulum is an essential organelle in the cell because it controls the functionality of proteins through the correct folding of their three-dimensional structure. Environmental stresses on plants alter protein homeostasis by causing ER-stress. Therefore, the UPR pathway is an essential signaling pathway to maintain and restore protein homeostasis at the cellular level. This review provides an overview of the UPR induction in cereals. In this work, two main tissues of the plant have been reviewed. In seedlings tissues, it has been described that during heat stress or chemical treatments inducing the UPR pathway, the UPR induction increases rapidly and reaches a peak. After, that maximum, the UPR induction decreases in the seedlings suggesting that the plant seems to use another strategy to alleviate to the ER-stress. In the grain compartment, the UPR is induced during its development under normal growth conditions and at higher levels during heat stress, suggesting a relation between processes involved in the accumulation of storage molecules in the seed and the involvement of UPR pathway. Therefore, in field crops such as cereals, cell signaling pathways in response to abiotic stresses in particularly complex plant compartments such as grain is little studied, although it is of unquestionable agronomic interest (harvestable part). Indeed, in cereals, the grain composition in starch but also in SSP is an important economic trait and it has been shown that under heat stress the quantity of SSP is decreased in the endosperm. Since these proteins are folded into the ER and that in response to ER-stress cells induce the UPR, we highlight that modulation of the UPR pathway in the ER organelle of the seed may modify the SSP composition and quantity determining seed quality and needs further investigation.
Also, since the effect of heat has already been investigated in plants and in cereals in particular; the involvement of the spliced form of bZIP69s as a messenger has not been completely proved. Evidence for systemic propagation of the UPR signals in plants has been shown by transduction of spliced bZIP60 from roots subjected to ER stress conditions to untreated tissues in Arabidopsis thaliana [65].
Systemic induction was attributed to cell-to-cell translocation of spliced bZIP60, either protein or mRNA or both, across plasmodesmata. This cell-to-cell mobility suggests that effectors of the UPR pathway could inform the whole plant of the physiological state induced by stresses, and could constitute a form of anticipatory protection. Further works need to be conducted to test the hypothesis that chemical or heat treatment might activate the UPR pathway (without triggering ERAD) leading to a better plant protection at cellular level from subsequent heat stress. Finally, the interaction between the two induction arms of the UPR pathway (bZIP28 x IRE1/bZIP60) constitutes a very poorly informed research axis in cereals, as well as the interactions with the cellular mechanisms linked to non-return phenomena (ERAD, autophagy, or apoptosis). These two arms co-exist and appear to have different functions in the response to heat stress that required be elucidated.
The authors declare no conflict of interest
This work has been supported by INRAe. The authors would like to thank Mahin Patel for reviewing the English of their manuscript

![]()

 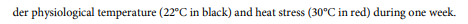 |
| Figure 1: Dynamic of UPR induction in cereals |
 |
| Figure 2: Overview of bZIP60s expression levels during heat treatment in cereals |
 |
| Figure 3: Overview of bZIP60s expression levels during DTT treatment in cereals |




































