Open Access
Research Article
Max Screen
ISSN: 2393-9060
Copyright: © 2024 Eduardo Festozo Vicente. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
During gestation, oxidative stress occurs physiologically and stressors such as endocrine alterations, maternal age, genetic problems, exposure to ethanol and malnutrition can con-tribute to this oxidative stress changing from physiological to non-physiological, impairing gestation, fetal development and reflecting on the adult life of the offspring. The objec-tive of this study was to evaluate the direct effects on sows and indirect effects on off-spring after adulthood of chronic exposure to ethanol and malnutrition while still in utero. The sows were divided into four groups: Control Group (C), ethanol (E), malnutrition (M) and ethanol and malnutrition (EM). Data were collected from the sows as soon as preg-nancy was confirmed and from the offspring from birth to the 60th day of life. The weight, food and water intake were evaluated in the sows and the weight, ferric reducing antioxidant power and FOX assays in the offspring. Maternal body weight showed a re-duction in group D from the 14th day of gestation. Group E decreased food intake from the first week of gestation and remained so until the end of the protocols. Regarding the hydration of the mother, groups E and EM showed decreased water intake from the first week of gestation and remained so until birth. Regarding the weight of the offspring, the measurement was performed on the 7th, 21st, 30th, 50th and 60th days of life, showing statistical difference in all groups when compared with each other or with group C until adulthood. Group EM showed a significant increase in FOX concentration compared to group M. Regarding the ethanol ingested by the mother, even in situations of malnutri-tion, during gestation it is capable of meeting the maternal and fetal energy demand in the short term. Therefore, it is in adulthood that there is greater production of peroxide lipids.
Keywords: malnutrition; ethanol; pregnancy; oxidative stress
Oxidative stress is defined as an imbalance between pro-oxidant and antioxidant factors. The term free radical refers to a highly reactive atom or molecule, which has an odd number of electrons in its last layer. When produced in excess by the body, they can cause cell and tissue damage, resulting in the phenomenon called oxidative stress [1].
During the normal gestational period, the maternal organism undergoes several changes, being heterogeneous in its physiological, metabolic and nutritional aspects that will directly reflect on the maternal and child health condition [2] but oxidative stress be-comes physiological during pregnancy due to a higher metabolic turnover and high resid-ual oxygen demands, with a greater need for antioxidant agents to maintain the balance between oxidant and antioxidant status [3].
The social consumption of ethanol has been increasing at an alarming rate among women worldwide, particularly among women of childbearing age (15 to 49 years) and pregnant women. Such behavior has been associated with maternal-fetal outcomes, such as intrauterine growth restriction, placental abruption, spontaneous abortions and congeni-tal anomalies such as fetal alcohol syndrome [4].
In women, the effect of ethanol consumption differs from that of men, even with equivalent intake for both. In women, the bioavailability of ethanol is greater due to its absorption capacity, higher proportion of fat mass, lower amount of body water and lower activity of alcohol dehydrogenase [5].
In the case of pregnancy, this scenario is aggravated, as the toxic effects can be po-tentiated both in the pregnant woman and in the fetus [6,7] which can cause direct tissue damage or indirect damage due to the presence of toxic metabolites responsible for stimu-lating the production of reactive species oxygen (ROS) and reduce antioxidant defenses [8,9], relating to increased oxidative stress, lipid peroxidation, reduced glutathione availa-bility and DNA changes [10, 11, 12, 13].
These molecular species are capable of inducing the maternal organism to react, through antioxidant systems, with the objective of limiting or inhibiting the production of these free radicals, repairing the damage caused to the cells or adapting the organism to this condition of oxidative [14, 15,16, 17]
Thus, to combat oxidative stress, the pregnant woman's body promotes increased production of antioxidants such as vitamin E, ceruloplasmin and erythrocyte thiols, releas-ing these substances into the bloodstream [18]. Studies show that increasing levels of an-tioxidants, such as vitamins A, C, E, β-carotene, and glutathione, are the best ways to combat ROS and its potential harm to the fetus [19].
Recent advances in research suggest that an individual's health begins to be de-fined during intrauterine development. The terms fetal programming or developmental programming refer to the factors that affect the growth and development of the individual and reflect on health in adult life (long term) [20].
In this sense, stressors during pregnancy, including maternal malnutrition, endo-crine changes such as insulin resistance, maternal age (young or advanced), maternal ge-netic or embryonic problems, alcohol consumption can profoundly compromise fetal postnatal development and have repercussions for life adult [21].
Authors concluded in a study that the chronic administration of alcohol in mal-nourished rats led to a 25% decrease in body weight, with protein loss, when compared to the control group [22]. In rats exposed to malnutrition in utero and evaluated in adulthood, it was possible to observe that they have metabolic dysfunctions and reduced life expec-tancy, as oxidative stress can promote DNA damage and accelerate cell aging [23].
It has also been described that ethanol contributes to a lower metabolization of nu-trients, due to the excessive production of reactive oxygen species, in addition to the liver changes caused by it, therefore, ethanol consumption can also contribute to nutritional deficiencies [24,25]
Based on these data, the aim of this study was to evaluate the direct effects on pregnant mothers and the indirect effects on adult offspring of chronic exposure to ethanol and malnutrition while still in the uterus, given the lack of studies with both associated risk factors.
For the study, it was used conditioned male and female Wistar rats from the Cen-tral Bioterium of Marília Medical School (FAMEMA). The animals were kept in a con-trolled temperature environment (23 ± 2ºC) and light-dark cycle (12 − 12h). All experi-mental procedures were approved by the FAMEMA Ethics Committee on Animal Exper-imentation, protocol nº 371/16, which are in accordance with the Ethical Principles on Animal Experimentation adopted by the Brazilian College of Animal Experimentation (COBEA).
Initially, 16 male rats were used only for mating, receiving water and free will and 32 females were used, 8 of them for each experimental group, they were:
Control Group (C) - received water ad libitum and standard commercial feed;
Ethanol (E) - received a daily dose of 10% v/v ethanol solution in drinking water during pregnancy and standard commercial feed;
Malnutrition (M) - received the standard commercial feed diet with a difference in reduction of 50% from the C group.
Ethanol and Malnutrition (EM) - received 10% v/v ethanol solution in the drinking water and were concomitantly received the standard commercial feed diet with a differ-ence in reduction of 50% from the C group.
The amount of standard commercial feed offered to groups M and EM was de-fined by a previous pilot experiment, based on the consumption of group C from the first day of pregnancy until delivery, which was 25g of standard commercial feed, becoming offered 50% less (12.5g) for groups M and EM.
The methodology described by [21] was adapted for the E and EM groups. Thus, before pregnancy, the concentration of ethanol in drinking water was gradually increased: in the first week, 2% v/v; second week, 5% v/v; and third week, 10% v/v. The rats con-tinued to receive a 10% v/v ethanol solution during the four days of male cohabitation and throughout the gestation period. Immediately after the offspring were born, ethanol con-sumption was restricted, as shown in Figure 1.
After this, mating was performed, in which two females were kept in the pres-ence of one male for four days. During this period, pregnancy was checked every morn-ing at the same time (8:00 am). Verification was performed by vaginal smear and ob-served under a Zeiss® optical microscope (10/0.25 x 6.3 mm magnification) to verify the presence of pregnancy-indicating factors, such as sperm heads and tails and the estrous phase. Confirmation of pregnancy was considered the first day of pregnancy, when fe-male rats weighing 250 to 300 g (80 to 120 days old) were selected and identified into the experimental groups described above, placed in individual cages. Male rats were discarded after mating, since their purpose was only reproductive.
Thus, 16 rat pups were kept to avoid future deaths and the consequent lack of rats for biochemical experiments, and from birth the pups of both groups received food and water ad libitum and weaning took place on the 21st day of postnatal life.
The selected animals were then kept in collective cages (four animals per cage) until the analysis of the experiment and were weighed at birth and weekly from birth to adulthood (10 weeks of age) on the 7th, 21st, 30th, 50th and 60th days of life. The ani- mals were submitted to the CO2 chamber and killed by exsanguination, followed by diaphragm rupture.
Determination of antioxidant capacity was evaluated in the plasma by ferric-reducing ability of plasma (FRAP) technique, from adult male adult offspring exposed to different protocols in utero. This technique is based on the ability of plasma to reduce fer- ric ions (Fe3+) in ferrous (Fe2+) in the presence of 2, 4, 6-tripyridyl-s-triazine (TPTZ) at acidic pH (3.6), forming a blue-col- ored Fe-TPTZ complex. For that, solutions were prepared using a 2.5 mL of FeCl3.6H2O (20 mmol L-1) addition onto a 25 mL acetate buffer (3.1g C2H3NaO2.3H2O, 16 mL L-1 C2H4O2) pH 3.6, and 2.5 mL TPTZ (10 mmol L-1, HCl 40 mmol L-1). The optical density (OD) of this solution was determined by means of absorption measurement at 593 nm on a Bio Tek spectropho- tometer (BioTek Instruments, Inc., USA) after 5 minutes of incubation at 37°C. After that, 10 µL of the sample and 30 µL of ul- trapure water were added to the initial solution, and the variation at OD was monitored for 8 minutes and registered each 30 se- OD was calculated for each sample. The values for antioxidant ca- pacity were calculated according to Benzie and Strain(22). Briefly, the calculations were performed by comparing the absor- bance values obtained from the tests and the absorbance values of Fe2+ solutions of known concentrations (from 100 to 1,000 µmol L-1). The resulting reference values for plasma antioxidant capacity was 612 – 1,634 µmol L-1 (mean value = 1,017; stan- dard deviation = 206).
Lipid peroxidation, which indicates the degree of oxidative stress of the biological environment, was estimated in male adult off- spring from different experimental groups by analyzing the consumption of ferrous oxidation-xylenol orange (FOX) reagent, according to the technique described by Jiang(23) and adapted for plasma and serum by Arab and Steghens (24). This method- ology is based on the oxidation of Fe2+ to Fe3+ in the presence of lipid hydroperoxides and formation of Fe3+ complexes with xylenol orange, generating a characteristic color, which can be measured by spectrophotometry at wavelength 560 nm. The as- say was performed in a solution containing 81% neat methanol; xylenol orange (100 µmol L-1), H2SO4 (25 mmol L-1), LB- HT-1 (40 mmol), and ferrous sulfate (250 µmol L-1), in which hydroperoxides were determined in plasma and aortic artery samples. Quantitation of peroxide levels was performed by comparison with a standard cumene peroxide curve (0, 0.5, 1.0, 2.0 and 4.0 µmol L-1).
The results were evaluated by comparing groups using a posteriori Tukey-Kramer test. In addition, results were expressed as mean and standard deviation (SD), even though the ethanol level data were analyzed by a nonparametric test. In all experi- ments, P< 0.05 was considered statistically significant.
The evolution of maternal body weight showed a gradual increase as a function of pregnancy and was not altered by exposure to ethanol or by association with malnutrition. However, malnutrition alone significantly reduced the evolution of maternal body weight from the 14th day of pregnancy forward (Table 1).
Since malnourished rats consume all rations, food intake in this condition and in association with ethanol is undetermined. The ethanol group significantly decreased food intake from the first week of pregnancy and remained like that until the end of the protocols (Table 2).
Malnutrition did not determine any change in fluid intake by animals throughout the gestational period. However, exposure to ethanol alone or in combination with malnutrition significantly decreased fluid intake from the first week of pregnancy and re- mained so until birth (Table 3).
As for the weight of the offspring, the measurement was made on the 7th, 21st, 30th, 50th and 60th days of life, showing statisti- cal difference in al groups when compared to each other or group C until adulthood, even when compared to the EM group, as can be seen in Figure 3.
As described in the methodology, the assessment of antioxidant capacity was evaluated in male offspring after adulthood ac- cording to their respective groups (C, E, M and EM) by the FRAP method. Malnutrition, as well as in utero exposure to ethanol, did not change the plasma antioxidant capacity of male offspring after adulthood when compared to other experimen- tal groups
In the offspring of male rats, when comparing nourished and malnourished animals without exposure to ethanol in utero, no statistically significant change was observed in FOX concentration. Likewise, exposure to ethanol, compared to animals that did not consume ethanol, showed no significant change in FOX concentration. However, exposure to ethanol associated with malnutrition showed a significant increase in FOX concentration compared to exposure to ethanol alone (Figure 5).
Pregnancy demands a huge process of adaptation in the woman's body from conception. The physiological adaptations that oc- cur throughout pregnancy aim at adapting the maternal-fetal complex, as well as childbirth. Insufficient quality and quantity of food in this phase may be associated with the multiple burden of malnutrition, characterized by the coexistence of nutritional deficiencies, such as anemia and vitamin A deficiency [27].
According to the European Society for Clinical Nutrition and Metabolism (ESPEN), malnutrition can be defined as a condition of the body in which, due to insufficient supply or incorrect absorption of nutrients, there are changes in body composition, physical and mental impairment and adverse effects on the outcome of an underlying disease [28].
In the present study, both mothers and offspring at different stages of life after birth were investigated. In relation to the weight of the matrix, the weight evolution of the pregnant rats showed that the M group presented less weight gain in relation to the C, E and EM groups. These results follow the assumption found by Ferrari, Gabrielli and Mello [29], who found that malnour- ished sows had less weight gain than the nourished group, or the same weight loss if the female was young (80 to 120 days old). In addition, Grinfeld [30] in his study, demonstrated similarities between the weight of the ethanol group and control mice af- ter the administration of ethanol by intragastric gavage from the 5th day of gestation.
Alcohol is obtained through the fermentation of foods that contain sugars, and provides 7.1 kcal/g, being considered a nutrient. Due to its energy value, it can meet the caloric needs of an individual and lead to overweight and even obesity, depending on the frequency and consumption [31, 32] this may explain the evolution weight for the EM group.
However, food intake was reduced in group E. This change can be explained by the fact that sows that consumed ethanol showed greater satiety, since alcohol consumption also provides many calories. The study by Silva et al. [12] presented the same result, when verifying the decrease in maternal food consumption throughout the pregnancy in the ethanol group in relation to the control.
The type of energy provided by alcohol is different from others as it is not able to be stored in the body. Alcohol has priority in metabolization, as it is highly toxic and the body discards it quickly, thus altering other metabolic pathways, including the lipid oxidation pathway, a fact that favors the increase of fat deposits mainly in the abdominal region [33].
As for the water intake of the sows, the C and M groups maintained a gradual increase throughout the gestational period, while the E and EM groups showed a significant reduction in water consumption when compared to the C and M group and re- mained so since the first week of gestation. gestation until the birth of the offspring
Junior [34] in his study, showed that pregnant rats that received ethanol by gavage showed irritability in addition to showing signs of drunkenness and delay in recovery. Such conditions contributed to delayed search for food and water, contributing to anorexia and dehydration by causing irritability of the gastric mucosa. When investigating these animals by necropsy, the au- thor did not observe signs of mechanical or chemical aggression to the mucous membranes in any of the cases. Therefore, it is believed that irritability due to alcohol intolerance and the state of drunkenness are factors that interfere, in an important way, in maternal nutrition, at least experimentally. In clinical studies, however, intolerance is not observed, as in these cases alcohol intake is voluntary.
It is known that hydration is essential for life. Women of childbearing age have less need for fluid intake than men due to lower mass and lower proportion of body water, but in specific cases the recommendations may change, which is the case of pregnan- cy, in which the recommendation may exceed 3 liters/ day [35]. The blood volume of pregnant women increases gradually between the 6th and 8th week of gestation and reaches a peak at the 32nd to 34th week of gestation, with water being the main component of human tissue, comprising 83% of the blood [36].
On the other hand, in cases where alcohol consumption occurs during pregnancy, there is an influence on the secretion of an- tidiuretic hormone, increasing the rate of urine production, consequently dehydration, in addition to changes in hemodynamic variables that contribute to fetal alcohol syndrome. (SAF), increased risk of miscarriage, increased chance of maternal mortality and psychological disorders such as depression and tendency to reject prenatal care [5].
As for the weight of the offspring, it was evaluated at different times, still babies (up to the 21st day), young adults (from 30 to 50 days) and adults (over 56 days) [37].
The M or EM group presented the highest low weight index from the 7th to the 60th day of age when compared to the C and E groups, therefore, the present study suggests that malnutrition is the most important factor for the occurrence of low weight and not exactly ethanol, as this isolated factor did not determine major differences in relation to group C
According to GRINFELD, [30] ethanol occupies the position of “cause” of low birth weight while Boggan et al. [36] point out that ethanol does not interfere with birth weight, however, it is capable of reducing body weight after the 28th postnatal day. Therefore, it is possible to state that alcohol is able to eventually supply the energy demand of the maternal organism, but not for the fetus/child, a result confirmed by several studies, which show less body weight gain from the birth of the offspring to adulthood. [39, 40; 41, 42].
It is believed that the harmful effects of ethanol can be identified even in utero, as it directly affects fetal growth by obstructing the transport of nutrients, with a decrease in the supply of oxygen, the substance promotes vasoconstriction of the placenta and umbilical cord vessels. With the decrease in the supply of oxygen and nutrients, there is an increase in oxidative stress in the pla- cental villi, with a decrease in the production of nitric oxide and, consequently, a decrease in blood flow [43].
Several studies show that ethanol is a determining factor for low birth weight in offspring [9,44] and, on the other hand, a study in humans, did not show significant differences between the body weight of newborns -Born to alcoholic mothers, however, were shorter at birth than the group not exposed to the substance [43].
It can also be suggested that the effects of ethanol can then be aggravated by malnutrition, as the toxicity associated with low caloric demand can influence the weight of the offspring. Thus, uterine malnutrition determined by low nutrient intake can be accentuated by the indirect action of alcohol, which reduces the ability to metabolize nutrients. When studying rats exposed to malnutrition and ethanol in utero, authors found that alcohol and malnutrition promote similar effects, both factors, alone or in combination, are able to negatively affect the increase in weight of the offspring at birth and in young life [45].
The toxic effects of alcohol in the body are mediated by oxidative stress through various mechanisms such as induction of oxidative damage, lipid peroxidation, cross-links and DNA strand breaks, and at the cellular level mitochondria are the main sources of reactive oxygen species (ROS ) [46].
Ethanol also impairs the cellular energy process, as it alters the transport and use of glucose, which is necessary for the develop- ment of the embryo's nervous system [47,48]. Alcohol-induced oxidative stress has been shown to be related to impairment of antioxidants, including superoxide dismutase (SOD), catalase (CA) and glutathione peroxidases (GPX) activities [49].
Free radicals attack carbohydrates, proteins, lipids and nucleic acids, whose damage can lead to cell death. The brain is the main target because it contains a high concentration of lipids and consumes large amounts of circulating oxygen, in addition to decreasing the endogenous antioxidant defenses of the offspring subjected to ethanol in utero, including SOD, CAT and GPX enzymes, all responsible for radical detoxification superoxide and hydroperoxides [50].
In the present study, it was used the FRAP method to analyze the antioxidant capacity of plasma, since a lower FRAP indicates a lower binding capacity of ferritin to iron and, consequently, a greater amount of free iron, capable of catalyzing the genera- tion of OH• radicals, through Fenton and Haber-Weiss reactions [51], it was found that there was no statistical difference be- tween the groups, not determining differences in the antioxidant capacity of adult rats exposed to ethanol or malnourished in utero or even in the association group. This fact can be explained by Ojeda et al. [42], who pointed out in their research that there may be a cellular adaptation of the adult offspring in relation to the antioxidant capacity of the plasma.
This adaptation can occur through three possible mechanisms: i) the first mechanism acts by preventing the formation of ROS by inhibiting the chain reactions characteristic of this process. Despite this, when there is the formation of free radicals, antioxi- dants are able to intercept these molecules, preventing the attack on lipids, amino acids of proteins and DNA, avoiding injuries and loss of cellular integrity [52]; ii) the second process is related to the removal of DNA molecule damage and the reconstitu- tion of damaged cell membranes. In response to the excessive production of ROS, the organism may adapt by increasing the synthesis of antioxidant enzymes [53].
In addition, another mechanism used by the body in the defense against free radicals is the non-enzymatic antioxidant system, also known as exogenous antioxidants, because they are, in most cases, from the diet, the consumption of these nutrients can improve the antioxidant defense and inhibit the formation of ROS [54]. The results referring to the FOX experiment indicated a statistically significant difference only for the EM group. This shows that exposure to ethanol alone is not a determining risk factor for oxidative stress. However, malnutrition can increase lipid peroxidation alone and in association with ethanol. Lipid peroxidation can be induced mainly by the enzyme NADPH oxidase, as this is one of the main sources of ROS in cells, being considered an essential factor in vascular dysfunction induced by ethanol consumption [55]. According to the results of Simpli- cio et al. [56] even acute ingestion of ethanol is capable of increasing the generation of ROS derived from NADPH oxidase. With regard to the malnourished group, the present study complements the work by Franco et al. [57], which demonstrates that maternal malnutrition is capable of causing endothelial dysfunction in the offspring, caused by increased activity of the NADPH oxidase enzyme. This fact explains the great difference between these results and those of the FOX experiment, which points to malnutrition as a more relevant risk factor than ethanol consumption.
Investigations addressing the effects of in utero malnutrition on ethanol exposure in humans are still very limited. Although it can be done, this search takes a long time before the results are available. In such situations, an animal model represents a ratio- nal alternative and, although it cannot be used to make unequivocal claims about effects in humans, it can significantly add to the body of evidence. Furthermore, food restriction in rats has proven useful as a model of human malnutrition, as these ani- mals share many characteristics of human malnutrition [58, 59].
This study provides important observations on prenatal ethanol exposure. This situation, regardless of in utero food malnutri- tion, led to a significant reduction in food and ethanol consumption during pregnancy. In contrast, significant changes in ma- ternal weight were observed only in the malnutrition group from the 14th day of pregnancy, as well as in the weight gain of the offspring. A lower plasma antioxidant capacity of the association group was identified when compared to the other groups. However, the results did not determine significant differences, while the production of lipid peroxides was increased in the asso- ciation group. Given these results, it is possible to state that maternal consumption of ethanol, even in situations of malnutri- tion, during pregnancy is able to meet maternal and fetal energy demand in the short term, nevertheless, it is in adulthood that occurs a higher production of lipid peroxides.

![]()

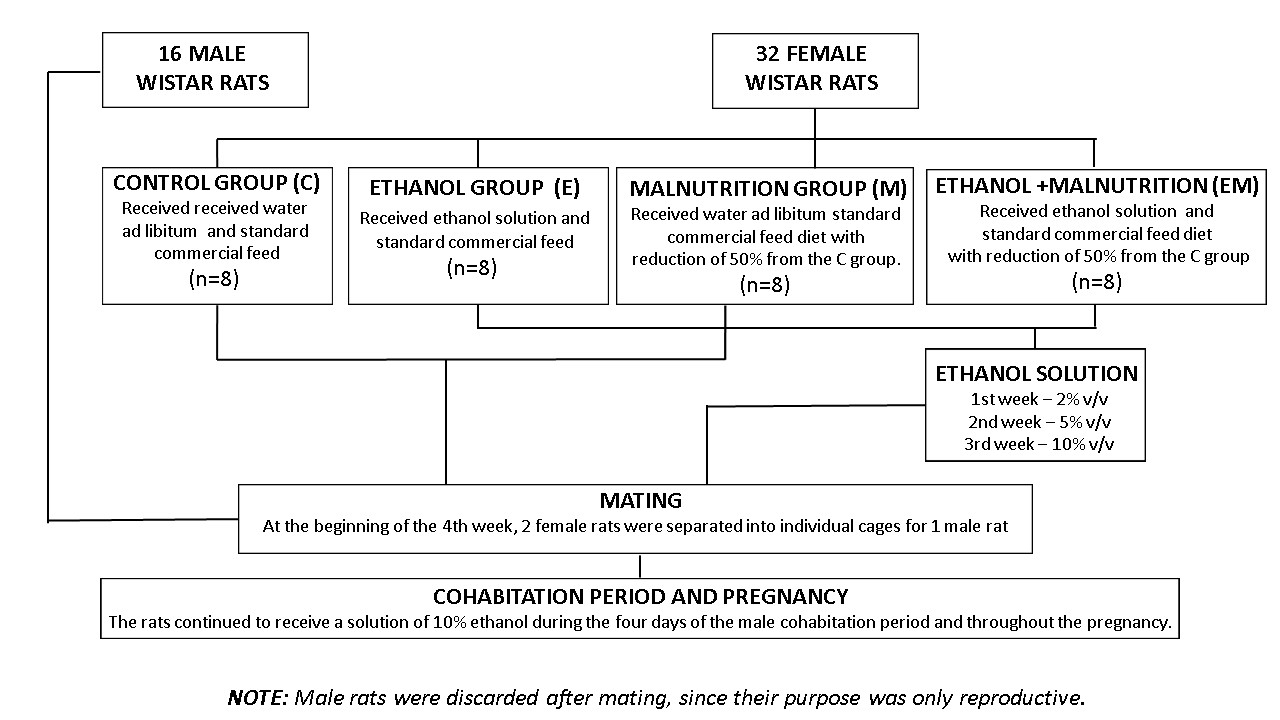 |
| Figure 1: Methodological strategy Source: elaborated by the authors (2023) |
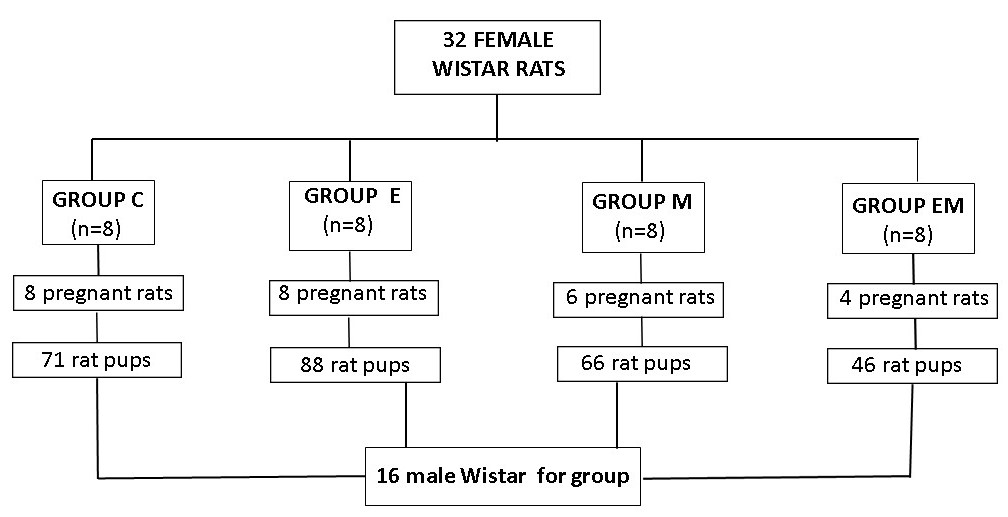 |
| Figure 2: Selection of puppies for research Source:elaborated by the authors (2023) |
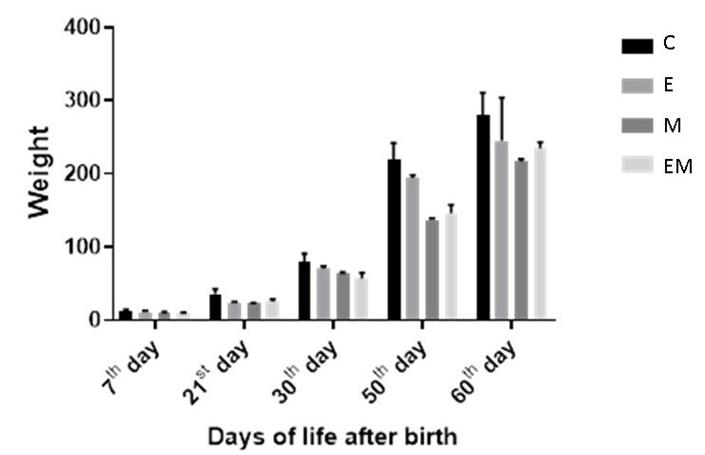 |
| Figure 3: Evolution of offspring weight from 7th to 60th day of life Note:values are means ± SEM. Number of animals per group = 8 |
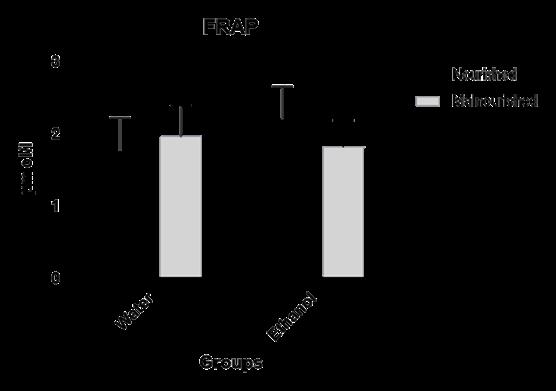 |
| Figure 4: Plasma antioxidant capacity determined by the FRAP method in adult male rats exposed to in utero malnutrition and
ethanol, alone and in association. Note:values are means ± SEM. Number of animals per group = 8 |
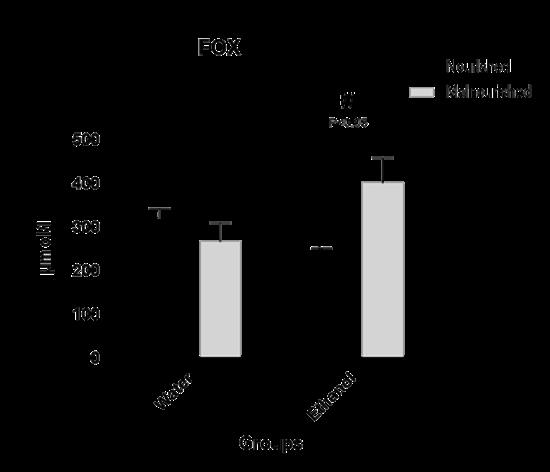 |
| Figure 5: Degree of oxidative stress in offspring of adult male rats exposed to in utero malnutrition and ethanol, alone and in combination. Note:values are means ± SEM. Number of animals per group = 8 |
Period |
Treatment groups |
P-value |
|||
C (n=5) |
M (n=5) |
E (n=5) |
EM (n=5) |
||
Day 0 of pregnancy |
266.2 ± 5.7 |
255.0 ± 35.0 |
267.5 ± 7.3 |
282.3 ± 9.3 |
> 0.05 |
7th day of pregnancy |
275.0 ± 6.7 |
239.3 ± 33.3 |
281.1 ± 6.3 |
257.3 ± 18.2 |
> 0.05 |
14th day of pregnancy |
295.0 ± 8.3 |
225.3 ± 28.6* |
290.5 ± 4.1 |
264.6 ± 20.5 |
< 0.05* |
21st day of pregnancy |
333.0 ± 11.3 |
243.3 ± 24.9* |
332.3 ± 9.2 |
299, 3 ± 24.1 |
< 0.05* |
Period |
Treatment groups |
P-value |
|||
C (n=5) |
M (n=5) |
E (n=5) |
EM (n=5) |
||
th |
23.8 ± 1.0 |
- |
15.3 ± 0.6 * |
- |
< 0.05* |
th |
24.7 ± 1.1 |
- |
16 , 6 ± 0.5 * |
- |
< 0.05* |
st |
26.0 ± 1.2 |
- |
19.0 ± 0.4 * |
- |
< 0.05* |
Period |
Treatment groups |
P-value |
|||
C (n=5) |
M (n=5) |
E (n=5) |
EM (n=5) |
||
th |
33.2 ± 2.2 |
30.3 ± 0.3 |
20.6 ± 2.4 * |
25.0 ± 2.2 * |
< 0.05* |
th |
41.7 ± 3.1 |
35.5 ± 0.5 |
29.5 ± 0.8 * |
31.5 ± 0.9 * |
< 0.05* |
st |
47.0 ± 1.2 |
40.6 ± 2.5 |
29.9 ± 0.8 * |
36.0 ± 3.4 * |
< 0.05* |




































