Open Access
Research Article
Max Screen
ISSN: 2393-9060
Copyright: © 2023 Ahmed Y Nassar. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
In this study, prepared acetylated bovine whey (ABW) was tested as a highly efficient hepatoprotective agent like that of the old drug N-acetyl cysteine (NAC) against the induction of iron overloading (IO) in a rat model. The probably prophylacting antioxdative characteristic features that were prominently executed on tissue content of reactive oxygen species (ROS), antioxidative ferritin, and proinflammatory IL-6 via the enhanced tissue protecting Nf-E2-related factor 2 (Nrf2) pathway. The reduction in macrophagic M2 CD163 and Kupffer M1 CD68 as well as apoptotic caspase 3 distribution denoted the efficacy of the targeting cellular prophylacting activity. In addition, the reduced abnormalities in the hepatocellular nuclei, cytoplasmic reticulum, mitochondrial degeneration as well as granular glycoproteins, and secondary lysosomal appearance promoted the more preferable usage of the natural product whey supplement than the risky NAC in IO cases
Keywords: Iron overload; acetylated bovine whey; N-acetyl cysteine; antioxidant; Nrf2
Iron chelating drugs are effective in reducing threats in iron overload patients [1]. The N-acetylcysteine (NAC) drug in vivo provides the amino acid cysteine, the main precursor for glutathione (GSH) biosynthesis. Dietary cysteine should be first activated to be absorbed in the intestinal epithelium, while oral NAC is passively transported [2]. Hence, NAC is a direct good cysteine provider for the tissue to get its required GSH. Since the homeostatic cysteine/cystine redox system is one of the most abundant in the systemic blood and body fluids, the corresponding redox system in the living tissues is the GSH/GSSG redox ratio [3]. In mammals, higher significantly elevated iron levels in the systemic blood, or serum ferritin (SF) which is an indicative marker of an iron overload condition, induce organ damage in the liver, heart, pancreas, thyroid, and central nervous system [4].
The main organ damage is mostly referred to an overproduction of reactive oxygen species (ROS) in the presence of such iron overload condition, the pathological conditions that are known as genetic iron overload or secondary syndromes including iron deposition resulting in cellular death, fibrosis and tissue injury or organ dysfunction or even tumarogenesis [4].
The hydroxyl radical is the most threatening toxic radical in ROS initiated by iron overload affecting the nucleic acid base 8-hydroxyguanine (8-OHG) that promotes tissue organ teratogenicity and carcinogenicity [5].
The oral-supplemented NAC has been observed to protect against ROS accumulation in the biological system by enhancing the antioxidant nuclear erythroid 2-related factor 2 (Nrf2) mRNA expression [6]. NAC acts by increasing the intracellular content of cysteine for GSH which scavenges free radicals. The interactions are counterbalanced by antioxidant enzymes, such as enzymatic transcriptions, that are regulated by genes containing antioxidant response elements (AREs). Such players as catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPX) via the Keap-1-Nrf2-ARE pathway [7]. Living mammalian cells have elaborate defense mechanisms for minimizing physiological stress. Nrf2 plays a crucial role in the cytoprotective response to oxidative stress and other cellular intoxications [8]. In unstressed conditions, the living tissue Nrf2 (t/2 is 20 m) content is maintained at low levels by its associated protein 1 (Keap1), an adaptor component of a Cullin 3-based ubiquitin E3 ligase complex, ubiquitin-proteasome degradation activity, and its components recycled [9]. Under oxidative stress and/or intoxication conditions, the specific cysteine sensors of Keep 1 are modified to inhibit the ubiquitination of Nrf2, which leads to the stabilization and accumulation of Nrf2 [10]. Such cytoplasmic accumulated Nrf2 translocates to the nucleus, where the (AREs) regions are ready to conjugate the signaling Nrf2 at the corresponding promoter sites for gene series expression of the antioxidant phase 2 enzymes [11]. Moreover, activated Nrf2 is highly effective for the maintenance of mitochondrial antioxidant defenses and redox homeostasis by disrupting mitochondrial thiol homeostasis of GSH metabolism [12]. Many synthesized cytoprotective proteins are up-regulated by Nrf2 including glutamate-cysteine ligase catalytic subunit (GCLC) and ligase of the regulatory subunit (GCLM) form a heterodimer in the biosynthesis of GSH [12]. The hemeoxygenase-1 (HO-1) enzyme that catalyzes the heme into the antioxidant biliverdin, the anti-inflammatory agent, is the Nrf2 target gene that protects from a variety of pathologies [13].
The glutathione S- transferases (GST) family members of cytosolic, mitochondrial and microsomal enzymes for the elimination of potentially harmful and toxic chemicals as conjugated with GSH are induced by Nrf2 activation [14]. Multidrug resistance-associated proteins (MrPs) of membranous efflux transporters acting from tissue organs to plasma to be excreted in urine or feces are up- -regulated by Nrf2. In addition, Nrf2 is ubiquitously expressed in animal tissues at different levels; in the kidney ˃ muscle ˃ lung ˃ heart ˃ liver, and brain [15]. Furthermore, Nrf2 inhibits the nucleotide-binding domain leucine rich repeat protein 3 (NLRP3) inflammosome activation in the human innate system [16].
Not only antioxidative features are initiated by NAC, but it can also act as a depicting chelator thiol and hydroxyl chelating sites to complex with metals [17]. In this aspect, some records have identified that NAC acted as a chelator of heavy metals in animal cell lines, one of which in vivo recorded that rats exposed to mercury then were treated with NAC and Zn mixtures prevented blood and liver mercury retention.
It had been reported that acute and chronic NAC treatment reduced the cellular concentration of Cu and Zn as well as modulated the homeostasis of these elements both in vitro and in vivo and accordingly affected the redox balance. The authors attracted attention to their observations that under conditions of limited trace element intake, homeostasis could be more susceptible to disturbance by chronic NAC intake. In addition, high doses of NAC inhibited the Zn-induced Nrf2 activation and limited the concomitant up-regulation of cellular GSH levels. In such areas of discrepancy NAC which protects against GSH depleting hepato and/or nephrotoxic xenobiotic under conditions of GSH deficiency [3], is typically ineffective in elevating GSH under normal conditions [18]. The case that had been verified by oral NAC administration to persons only increased blood cell GSH levels in individuals who had unusually low GSH levels before treatment [19]. By the way, NAC as a nutritional supplement, is found in some fruits and vegetables in small amounts.
Noticeably, as an oral therapeutic prodrug, NAC should be more than 1200 mg/day in humans [20] because NAC provides the biological systems with cysteine for denovo GSH biosynthesis in severely depleted conditions. Its biological half-life is about 6 hours and clearance in renal and non-renal with side effects of nausea, vomiting and diarrhea [21].
In this area of negative NAC limitations; supplements may cause mild digestive upset in infants or allergic reactions. Children, people with bleeding disorders, and those with kidney diseases should avoid taking NAC.
Unfortunately, Schwalfenberg [22] in his alarming against chronic utilization of NAC, claimed that high plasma cysteine levels may indicate poor clinical outcomes in patients with acute stroke; possible involvement of hydrogen sulfide
Moreover, the high-dose of oral NAC modulates inflammation in cysteine fibrosis. It has been reported that the concentration of GSH in infants' RBCs is a heritable trait. In addition, Sawamoto et al [23] on chronic doses of NAC, found that L-cysteine-induced brain damage in experimental animals have presented a neurotoxic activity of L-cysteine interaction with glutamate. Also, Paschalis et al [24] verified that NAC supplementation is only valid in cases with low levels of GSH.
Anyhow, therapeutic NAC has special precautions and warnings as it should only be used in pregnant women when medically needed, it doesn’t use in people who are allergic to acetylcysteine, and it might cause bronchospasm in people with asthma if inhaled or taken by mouth or through a tube in the windpipe, NAC might slow blood clotting. There is concern that NAC might increase the risk of bruising and bleeding in people with bleeding disorders and during or after surgery so it should be stopped at least 2 weeks before scheduled surgery [25].
Recently, Nassar et al [26] have detected that oral acetylated whey exhibits potent antioxidant and anti-inflammatory characteristics to protect the spleen in iron-overloaded rats
Whey, which is the liquid part of the cheese-making industry made from bovine dairy milk, contains about half the solids of milk, especially proteins with different physical, chemical, and nutritional characteristics as well as several biological effectors. In the last decades, several researchers have been concerned with such peptides and their digested hydrolysates as therapeutic potentials [27]. The activity of such peptides is based on their amino acid composition (from 2 to 20 residues) and sequence. Antioxidative peptides from whey exert their effect by intracellular conversion of cysteine into GSH, which keeps cells safe from ROS by inducing genes. In general, the antioxidative peptides are closely related to the peptidomics composition and hydrophobicity [28]. Tyr, Trp, Met, Lys, Cys, and His are examples of amino acids with antioxidative activity. In addition, Rajapakse et al [29] proposed that the antioxidative activity of His-containing peptides is related to the hydrogen-donating lipid peroxyl radical trapping and/or the ionchelating ability of the imidazole group. Differentially, the SH group of cysteine has its own specific direct interaction with radicals. The hydrolysate of whey protein by alcalase enhanced the antioxidant activity in the liposome model system [30]. On the other hand, whey protein-derived peptides obtained from proteolytic digestion showed considerable binding activity with cations such as Ca2+, Fe2+, and Zn2+ [31]. Tripeptide Tyr-Asp-Thr was identified in the hydrolysate, where the major binding sites included the oxygen atom of the carboxyl group and the nitrogen of the amino or imino groups.
In this study, the predicted prophylacting NAC might be acted as an exogenous provider of derived intracellular cysteine for renewal of GSH that increased significantly the cell antioxidative Nrf2. On the same level of action, in vivo, ABW within cellular physiological requirements for synthesizing the intracellular GSH from cysteine or even other metabolized amino acids has been performed for such a pathway of action [12].
This study has been designed to use an afforded whey supplement as a natural product to be an efficient alternative competitively potent component exhibiting antioxidative and probable valuable metal chelating capacity like NAC or more in iron overloaded living model without harmful or probable threatening bi-side effects.
Forty - eight rats (100-150 g B.W.) under standard laboratory conditions were incubated and thoroughly selected for this study. They were equally divided into four categories; G0 for the healthy normal negative control, G1 for induced iron overloading positive control, G2 induced iron overloading simultaneously prophylactic with N- acetylcysteine (NAC) drug, and G3 induced iron overloading with simultaneously tested prophylacting supplemented acetylated bovine whey (ABW). Roat of adminered scheduled doses for iron overloading by iron dextran as well as treating NAC or prophylacting tested bovine whey ABW in our laboratory have been recently published [26].
Livers of all rats tested groups were singly characterized and excised to be washed with distilled water to get rid of remaining blood, then thoroughly dried by blotting filter paper. The dissected liver samples of each rat were assigned into four patterns of tissue samples
a- Part to be homogenized in phosphate buffer saline (PBS) and stored at -80oc for biochemical ELISA assay.
b- Part to be directly used in electrophorytically tested Western blotting assay.
c- Part to be fixed in 10% neutral buffered formalin for histopathological and immunohistochemical investigations.
d- Part to be in glutaraldehyde fixation form for electronic microscope examination.
In the aliquot of hepatic tissue homogenate that was considered for determining hepatic reactive oxygen species (O2- and H2O2), macrophagic Interleukin-6 and the synthesized reducing ferritin by ELISA kits (of INNOVA Biotech Company, China, Cat. NO.; In-Ra 1426, In-Ra 0688 and In- Ra 1156 respectively) according to their protocols.
Tissues from the rats under experiment (3mm3) were lysed with ice-cold RIPA buffer (50mM Tris-Cl (pH 7.6), 5 mM EDTA, 150mM NaCl, 0.5 % NP-40, and 0.5% Triton -X-100) containing 1 µg/ ml leupeptin and aprotinin, and 0.5 mM PMSF. Lysates were centrifuged at 2,500 rpm for 10 min at 4°C.
Protein concentrations were measured by Bradford assay. Thirty µg protein aliquots were separated by SDS PAGE using 10 % gels and then transferred to nitrocellulose membranes. Membranes were blocked with 5 % skim milk and probed with primary antibodies (anti Nrf2 IgG and β-actin 1:1000) overnight at 4°C.
Membranes were then incubated with HRP- conjugated secondary antibody (1:5000) for 1 h at room temperature. Detection was performed using the ECL substrate. Band densities were measured by Image J software and normalized to the control then equalized to the corresponding actin band.
In Paris, the first documented case took place on April 12, 1829 when a mastectomy was performed by French surgeon Jules Germin Cloquet (1790-1883 AD). In the United States, the technique was applied initially on June 30, 1836, in a case of dental surgery, performed by Dr. Harwood, in Boston. In England, Prof. John Elliotson (1791-1868 AD) was the first to introduce this novel approach by inserting a seton into a patient’s neck [14].
Deposited iron in hepatic tissue was performed by fixating dissected tissues in 10 % buffered formaldehyde solution for one day. The specimens were embedded in paraffin wax and 5 µm sections were prepared for histological staining (Kiernan, 1999). Prussian blue (PPB) staining was achieved for the detection of iron deposition in the hepatic tissue specifically non-hem iron "hemosiderin (Kiernan, 1999), and investigated under LM at 100 x magnification. Hҩ E stained hepatic slides were examined at 100 x, 400 x and 1000 x magnifications. Sections from rats liver portal area (PA) were stained by van grieson red color stain for detection of hepatic collagen formations, vesioualizedat 400 x magnification.
Full-thickness hepatic tissue slices were fixed in 4% glutareldehyde for 4 h at 4 0c, cut into semi-thin sections (0.5 – 1 m thick), and stained with toluidine blue. Using the transmission electron microscope, ultrathin sections (50- 80 nm) were cut from selected portions of semithin sections, contrasted with uranyl acetate and lead citrate, analyzed using (JEM-100 CXII AKishima, Tokyo Japan) and photographed at 80 Kv in E.M. Unit of Assiut University (Bozzola & Russell, 1999).
The primary antibodies for the macrophagic heamoglobin – hoptoglobin complex scavenger, anti-CD163 (Mouse monoclonal antibody Abcam Cat. No; ab 156769) and the kupher cell marker anti-CD68, Dako Agilent Cat. No; IR 609) as well as anti- caspase. Paraffin-embedded tissue sections of 3-5 µm thickness were first deparaffinized with xylene and then hydrated in graded ethanol solution and heated in citrate buffer (pH 6.0) for 5 min. Next, the sections were blocked with 5 % bovine serum in PBS for 2 h. The incubated slides overnight at 40c with primary antibodies at dilution 1:100. The slices were rinsed with PBS for 10 min. in a solution of 2% diaminobenzidine (DAP). Sections were counterstained with hematoxylin, dehydrated, cleared in xylene and then cover slipped for LM examination.
Data were presented as mean ±S.D. the assayed markers; ROS, IL-6, ferritin concentration, and Nrf2 were compared between the four different studied groups G0, G1, G2, and G3 using one-way analysis (ANOVA).
As it is presented in Figure 1 where the iron overload induction on the exposed rats, showed the hepatic tissue concentration of ROS was significantly elevated. The prophylacting agents NAC and AWP restored the levels similarly to the normal healthy attitude.
The results denoted that iron intoxication (Fe) significantly increased the inflammatory response compared to the healthy control group G0 (++P< 0.01). While the concentration was relatively sustained as a pro-inflammatory marker in association with either NAC or AWP in iron overloading induction (Figure 2).
As presented in (Figure 3), the results showed that iron overloading in the animal under experiment (G1) significantly elevated the heap tic tissue ferritin concentration (+++P< 0.01) compared to the healthy control group G0. The association of NAC enhanced such elevation while orally supplemented AWP was the same what reduced than that of NAC
Results showed Nrf2 antioxidative gene was dramatically reduced by iron overloading (Figure 4). The normal healthy level was restored by the action of NAC or AWP tested prophylacting agents
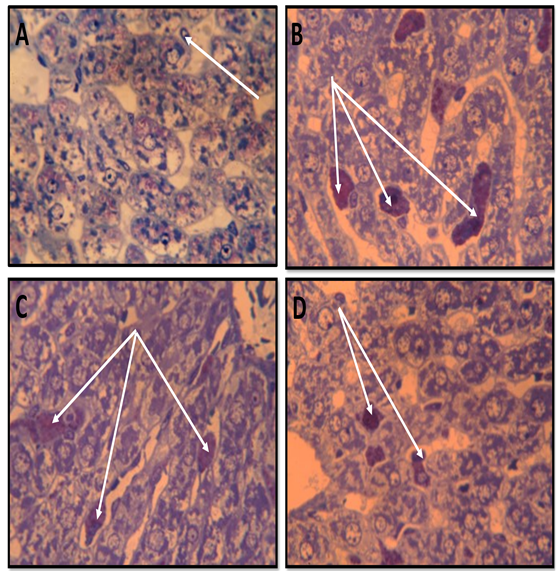
The iron distribution in the overloaded rats without treatment (G1) compared to the normal healthy (G0). the lowering prophylacting efficacy on such enriched iron intoxication was detected in NAC treated group (G2) and AWP (G3) treated group (presented in Figure 5 A, B, C, and D) via Prussian blue staining sections. Moreover, toluidine blue staining of other sections has verified the same features of hepatocytic iron intoxication of nuclei and blood sinusoids lined by Kupffer cells and reduced iron intoxication by active tested NAC or AWP supplement (Figure 6).
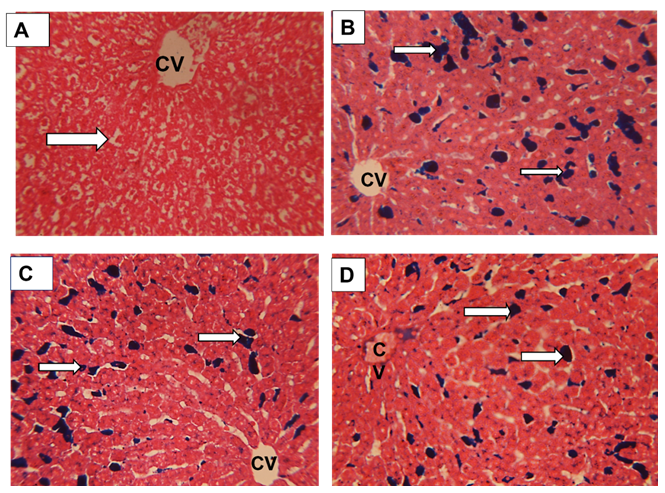
The normal rat liver (G0) showed normal healyhu structure in the central vein (CV) and portal area (PA) regions (Figure 7 A, B). The iron overloading (G1) presented enriched Kupffer cells (KCs) of blood sinusoids engorged with the iron (Figure 7 C, D). The prophylacted NAC group (G2) and the supplemented ABW (G3) showed lesser distribution (Figure 7 E and F as well as G and H respectively).
The stained collagen in the iron-overloaded animals (G1) was prominently increased compared to the healthy normal ones. The prophylacting NAC or AWP supplement reduced such synthesis. The AWP seemed more improving one comparing to that of NAC (Figure 8).
The hepatic markers CD 68 and CD 163 were increasing by iron intoxication (G1 ) when compared to the normal healthy group (G0). Such an increase was somewhat reduced by pretreatment of NAC (G2 ) or ABW supplement (G3 ) when compared to ironoverloaded rats (Figure 9 and 10 respectively).
The healthy normal (G0) slightly responded cases showed an intensive reaction. Pretreatment with NAC (G2) slightly reduced the response while ABW pretreatment (G3) dramatically reduced the staining (Figure 11).
Effect of Iron Overloading on the Hepatic Structure Using E.M. InvestigationElectron micrographs of healthy normal control rats (G0) under examination showed normal hepatocytes with the euchromatic nucleus, well-developed rER and mitochondria, lipid droplets, and KCs lining the blood sinosoides. The iron overloading (G1) showed hepatocytes with irregular heterochromatic nuclei surrounded by many vacuoles, KCs enriched with heavy iron overload, and irregular secondary lysosomes. Plenty of amalgamated mitochondria with each other and ill-defined cristae and electron-dense matrix (Figure 12 A and B as well as C and D respectively).
The overloaded pretreated with NAC (G2) group of rats under experimentation showed maitanance of crowded KCs with precipitated iron in the hepatic tissue. Also, the presence of increased secondary lysosomes and smaller size and amalgamated mitochondria were preminantly characterized (Figure 13 A and B). The overloaded pretreated with the ABW supplemented rats (G3) showed a smaller size of KCs in the hepatic tissue with the lower sectional distribution of secondary lysosomes. The mitochondria appearance was appearing in a more improved and healthy appearance in structure (Figure 13 C and D).
Basically, our health and diet are one of the considered pathways to prevent diseases, one of which is the iron overloading that could be prevented by whey, due to its high organic beneficial chemical nutritional, and biological properties. The putative biological and physiological effects of α- La, β- Lg, Lf, lactoperoxidase, immunoglobulins, glycol macro peptide (GMP), protease peptone, and a variety of growth factors as well as cysteine the building block of GSH, as well as the dietary antioxidant [32]. In addition, some digested whey proteins exhibited inhibitors as natural products acting against a wide array of tumors via an apoptosis like event [33]. The induced iron overloading in the experimented rats has been executed in this study.
The observed heavily stained iron widely distributed in the liver slices of the overload G1 was locally predicted in hepatocytes, sinusoidal KCs, and endothelial cells as it was observed [34]. Such tissue iron enrichment especially in KCs may be attributed to provoked macrophagic immunological response against induced iron intoxication. This G1 enriched hepatic tissue iron compared to the healthy normal (control group G0) was associated with significantly elevated levels of assayed ROS as O2- and H2O2 that should be attributed to a considerable increase in the elevated percentage of free iron in such iron-overloaded sections [35].
These threatening indices of oxidative stress promoted the predicted pro-inflammatory cytokine IL-6 biosynthesis [36]. Noticeably, these iron intoxication biomarkers were going side by side with those of frankly detected histological features of characteristic hepatic injury as well as cellular apoptosis [37]. The detected reduction in iron stain distribution in the treated groups; (G2 and G3) was mostly denoting a similar powerful metal chelating activity where the powerful metal chelating binding SH and neighboring OH residues in NAC structure [38] or SH and NH2 in different digested whey oligopeptides [39] that were afforded.
The macrophagic immune response that was prominently elevated by NAC, resulted in the assayed increased antioxidant ferritin synthesis [40] in the chronically iron intoxicated livers of G2. In this aspect, Wallace [41] has stated that chronically used NAC may enhance macrophagic receptor formation for further iron-importing from the surroundings and consequently, this imported iron was rapidly complexed with the antioxidant ferritin apoprotein. The previously recorded IL-6 as an acute phase protein enhances ferritin synthesis after receptor-mediated endocytosis of transferrin-binding iron (TBI) and other iron concomitantly imported. In this area of interest, Sebastiani et al [42] claimed that increments of macrophagic iron may be enhanced by the IL-6-hepcidin pathway. Our results denoted that either NAC or tested ABW exhibits positive pro-inflammatory responses in IO cases. NAC molecular mechanisms are known to include anti-inflammatory actions in intoxicated hepatic tissue. Noticeably, the tested whey supplement in this study has resulted in a closely related effect that looks like that of NAC whatever for assayed IL-6 or the hepatic ferritin content, denoting that the whey peptidomics action may participate in the preparation of the macrophagic receptors and other cellular immune response in hepatic iron overload (the task that should be approached in another study). Interestingly, the hydrolysis of α-Lactalbumin and β-Lactoglobulin by digestive enzymes may promote specific immune response through modulation of splenocyte proliferation and cytokine section as was observed by Brandelli et al [43] and recently by supplemented ABW to the induced iron intoxicated animals [26]. In addition, the trypsin action resulted in peptidomics fractions with immunoenhancing effects on proliferation and phagocytic activities on macrophage like cells [44]. The increased free radicals may promote the accelerated reduction of cellular endogenous GSH especially in mitochondria [12], the action that efficiently requires renewal of GSH via the genetic Nrf2 pathway. The predicted cascade reaction has been verified by the herein applied Western blot identification of Nrf2, where in a previous study the NAC hepatoprotection against intoxicating agents was possibly performed via the Nrf2-HO-1 pathway [13]. The tested competitively supplemented ABW to that of NAC treatment deduced similar antioxidative Nrf2 activities in the iron-overloaded livers were rationally accepted because some protein hydrolysates via regulation of GSH and other thiol modification have been thoroughly studied and confirmed [45]. Generally, the cytotoxic hepatoprotection via the Nrf2 mechanism prominently promotes genes of the anti-inflammatory factors including the effector (PPAR-ɣ) that enhances the activation of M2-macrophagic gene which herein immunologically detected as CD163 and specifically CD68 of KCs. This peroxisome proliferator-activated gamma (PPAR-ɣ) is a lipid sensor in the affected tissue via regulating the transcriptional gene expression involved in hepatic inflammations. Considering the apoptotic immunostained caspase-3 identified in these induced iron overloads hepatic macrophagic tissue cells appeared as intrinsic mitochondrial associating cellular pathways that respond to the initiated oxidative stress [37].
The tested NAC as well as the ABW mostly acted as antioxidants and thiol reductants prevented such apoptotic cellular death
In this approach, Zhang et al [46] have recently investigated the effects of exogenous antioxidant NAC on tumor formation and growth using transcription analysis and found that the transmembrane Bax Inhibitor Motif Containing-1(TMBIM1) significantly upregulated even in low NAC concentrations. Noticeably, the Van Gieson stain in the tested sections for collagen showed in a portal or periportal area considerable sustained deposition on treatment with NAC. The authors have differentiated between the exogenous and endogenous renewal of intracellular GSH where periportal collagen type 1 and 3 and matured elastic fibers initiated by acute or chronically exogenous GSH supplements probably promote cirrhosis [47] or even tumors. The used peptidomics ABW or even amino acids probably appreciated antioxidants against liver injury, the suggestion of the endogenous physiologic regulatory agents for physiological renewal of intracellular GSH
The digested whey protein fractions that had been characterized from α-lactalbumin or from β-lactoglobulin [43] exhibiting antioxidative effects were suggested to involve inhibitory characteristic actions against hepatic lipid peroxidation and scavenging of free radicals via specific cellular genes [48].
In this aspect, potent antioxidative amino acids could be involved in the digested whey fractions especially those with aromatic residues that can donate protons to electron deficient radical scavenging characteristics of the amino acid residues. For instance, the antioxidative activity of His-containing peptides relates to the hydrogen-donating lipid peroxyl radical trapping and/or the metal ion-chelating ability of the imidazole group. Moreover, the thiol (-SH) of cysteine exhibits specific antioxidant action related to its direct interaction with radicals [49].
Formerly, the antioxidant capacity assay technique by using 2,2'-diphenyl-1-picrlyhydrazyl for detection of both electron transferrin and hydrogen atom transfer concomitant with ferric ion reduction has been rationally considered [43].
Practically, specified enzymatic orally digested whey supplement in laboratory tested animals resulted in the decrease of creatine kinase, and lactate dehydrogenase in vivo biomarkers of oxidative stress and tissue damage. Moreover, the efficacy of whey protein derived peptides for binding cations is well-known as one of the best tested iron peptide complexes. The net charge, the side chain length, and functional groups of the amino acids and peptides are direct determinants of the extent of complex formation with iron. The authors recognized; the primary carboxyl groups, ξ-amino nitrogen of lysine, and the guanidine nitrogen of arginine are involved in iron-peptide bonding as well as glycine and proline. Others observed that glutamic and aspartic and their carboxylic groups were among the main-iron-binding complexes
Conclusively, because efficient oral therapeutic NAC should be more than 1200 mg/day in humans and stays for about 6 hours of clearance in renal and nonrenal associating nausea, vomiting, and diarrhea [22]. The concentration and duration can modulate hepatic cysteine fibrosis or even carcinoma and tumor growth via induction of TMBIM1 as exogenous GSH cellular renewal in injured liver tissue [46]. The edible digested natural product of whey showed efficient chelating activity against induced iron overload similar to that of NAC as well as the antioxidative gene Nrf2 up-regulating pathway. Such activated Nrf2 predicted chemoprotective effects against mitochondrial damage (as was observed in the electro scanned photos). In addition, such activated Nrf2 regulators mostly balanced the mitochondrial hepatic ROS by promoting detoxification of accumulated peroxides associated with induction of iron overloading via replenishment of the depleted GSH.
Concomitantly, this predicted AWP promoted the induced highly toxic iron to be more complex as reducing ferritin in hepatic tissue and macrophages, especially KCs via receptor signaling. Moreover, oral-supplemented whey is mostly predicted as the best provider of the hepatic tissue precursors for physiological pathways to the endogenous renewal of GSH without enhancement or deposited fibrosis. In addition, the probable whey derived amino acids Tyr, Trp, Met, Lys, Cys, and His are predicted examples of amino acids that exhibit antioxidant activity could be provided. Hence, this study may appreciate the use of oral whey supplement to be the more preferable subsidizing hepatoprotective instead of the chronically risk old NAC drug, acting against iron overloading features in intact living animals.

![]()

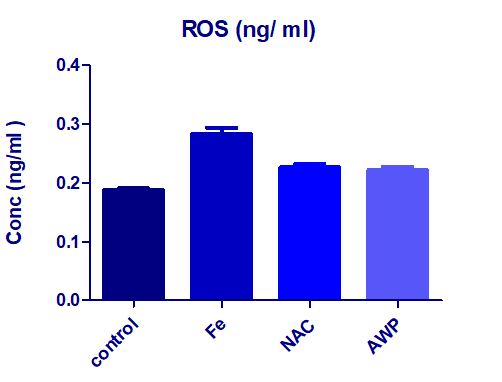 |
| Figure 1: Effects of NAC and AWP (G2 and G3) on the hepatic tissue ROS concentration presented as mean ± S.D. ++P< 0.05 compared to iron overloaded group G1 (Fe) deduced by one away ANOVA (+++ significant when compared to healthy control group G0). |
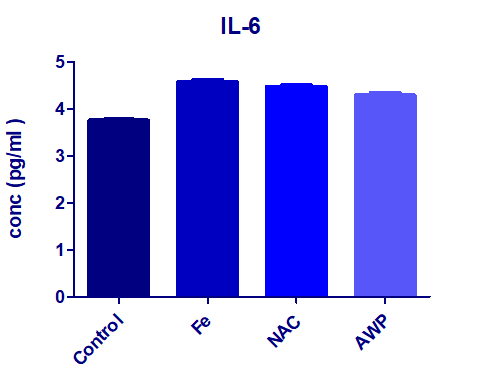 |
| Figure 2: : Effect of iron overloading on the pro-inflammatory response marker IL-6 presented as mean ± S.D., showed significant increase (++P< 0.01) compared the level of healthy normal control group G0. In presence of NAC or AWP (G2 or G3) as prophylacting agents did not show considerable change when compared to the non trated group G1. |
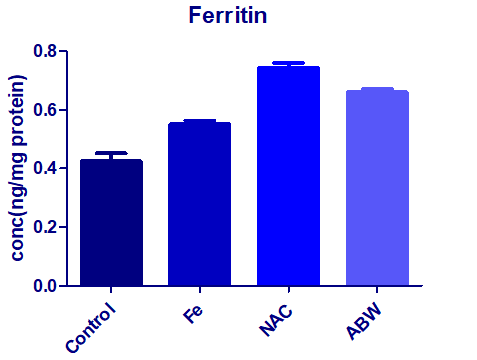 |
| Figure 3: Effect of iron overloading (G1) on the hepatic ferritin concentration as mean ± S.D. (+++P< 0.01), comparing to the normal control G0. The associated treatment with NAC (G2) or with the supplemented AWP (G3). |
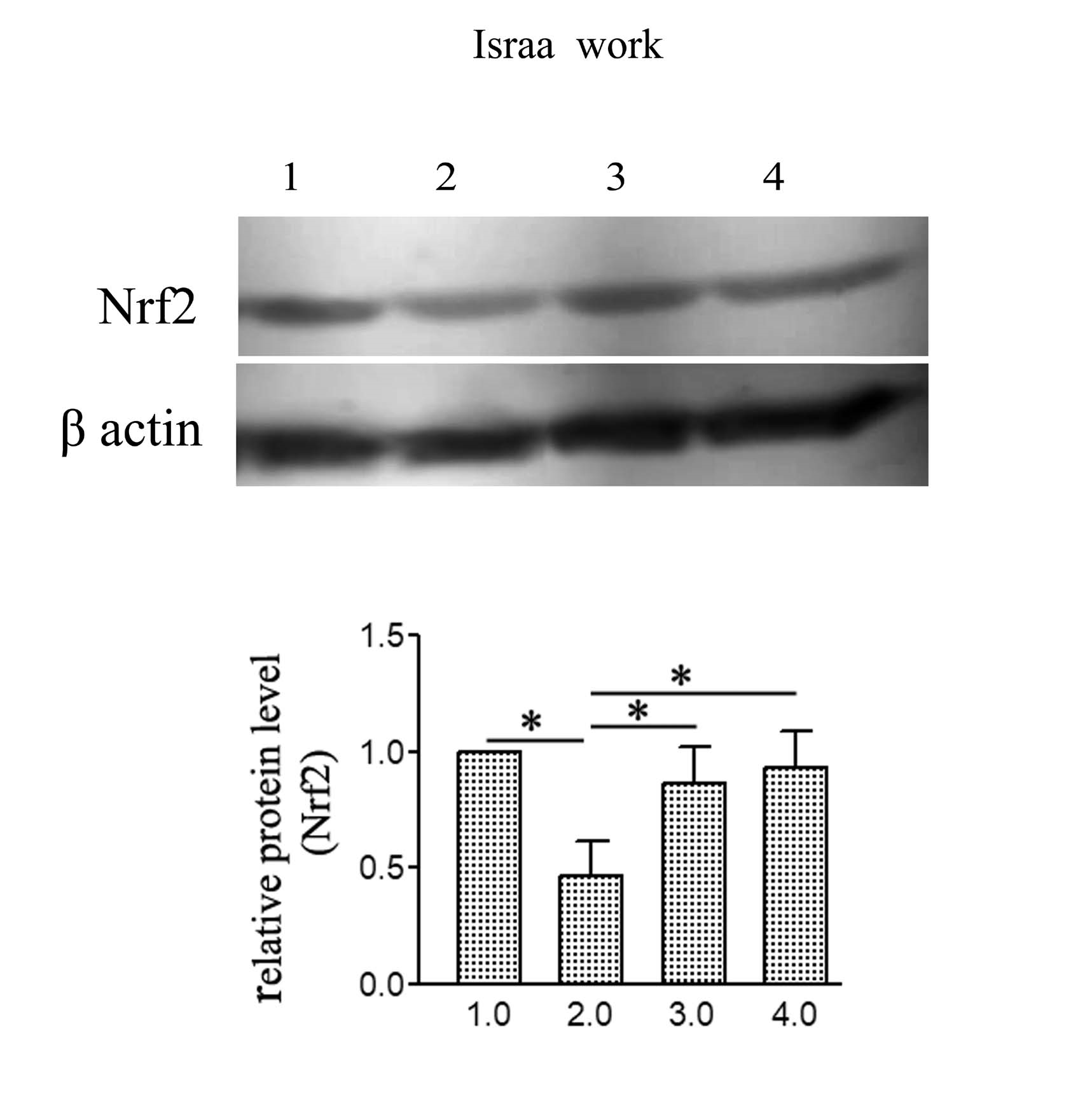 |
| Figure 4: Impact of NAC or AWP prophylacting agents on the protein bands detected with ECL in the iron overloaded rats. The presented values; mean ± S.D. (+P< 0.01) compared to iron intoxicated group (G1), while statistical analysis and comparisons by using one-way ANOVA. |
 |
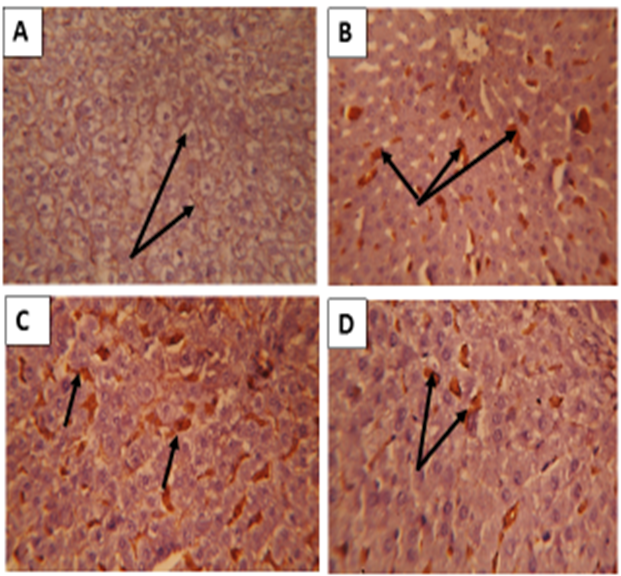
| Figure 5: Sections in rat liver stained by Prussian blue for demonstration of iron showing; (A) Healthy control group showing no deposition of iron(arrow) .( B) Iron overload intoxicated group showing high population of sinusoidal Von kupffer cells over loaded with iron( arrows) .(C) N-acetyl Cysteine treated intoxicated group showing some decrease in iron loaded Kupffer cells (arrows) . (D) Whey- Acetylated peptide treated intoxicated showing evident decrease of iron - loaded Kupffer cells ( arrows) .X400 |
 |
| Figure 6: Semithin sections in rat liver stained by Toluidine blue showing ; ( A) healthy control group with hepatocytes having normal active nuclei ( thin black arrows) and blood sinusoids lined by normal squamous cells and Kupffer cells free of any iron deposition( whit arrow). (.B) Iron overload intoxicated group showing numerous iron loaded Kupffer cells (white arrows). (C)&(D) N-acetyl Cysteine and WheyAcetylated peptide treated groups showing that the iron loaded cells (white arrows) appeared more or less similar to the intoxicated group with normal hepatocyte . X1000 |
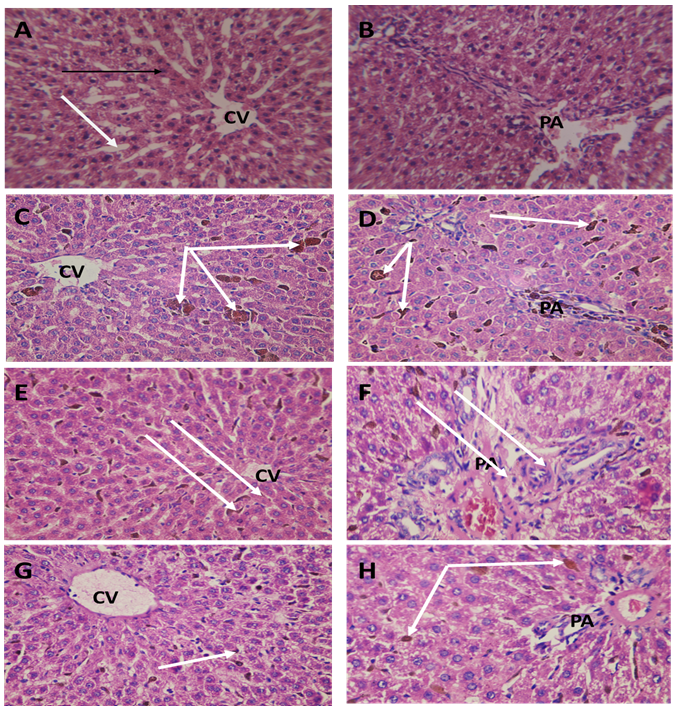 |
| Figure 7: Sections in the rat liver from rat central vein(CV) and portal area(PA) regions stained with HX&E stain to show:. (A&B) Healthy control group G0 showing the normal structure of the liver with thin walled blood sinusoids (white arrow) and normal hepatocytes cell cords, ( C&D) Iron overload intoxicated group G1 showing the Kupffer cells of the blood sinusoids appeared engorged with iron( white arrow) . (E&F) N-acetyl Cysteine treated intoxicated group G2 showing the Kupffer cells of the blood sinusoids appeared engorged with iron but lesser than the iron overload group. (.G&H) Whey- Acetylated peptide treated intoxicated group G3 showing the Kupffer cells of the blood sinusoids appeared engorged with iron (white arrow) but lesser than the iron overload and N-Acetyl Cysteine treated groups. X400 |
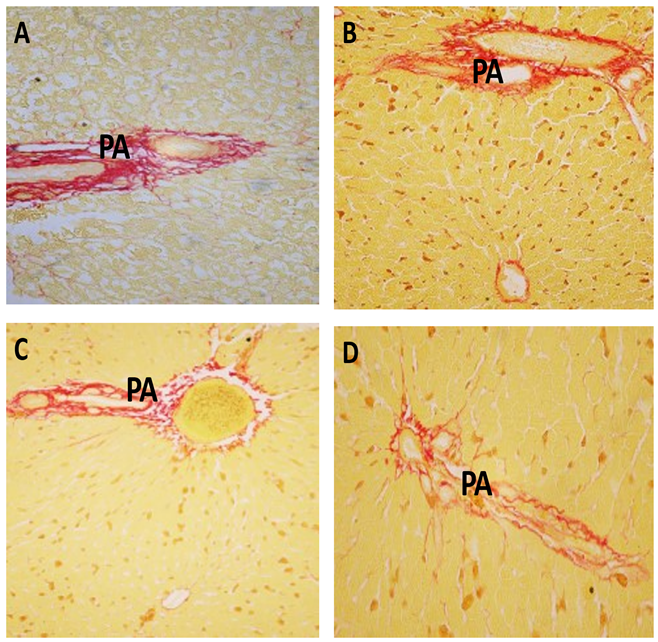 |
| Figure 8: Sections from rat liver portal area(PA) stained by Van Gieson stain ( red color) for collagen to show :(A) Healthy control group showing normal collagen content in the portal area (PA) . (B) Iron overload intoxicated group showing an increase in collagen deposition in the portal area(PA) . (C) N-acetyl Cysteine treated intoxicated group showing moderate decrease in collagen content. (D) Whey- Acetylated peptide treated intoxicated showing marked decrease of collagen content. (X400) |
 |
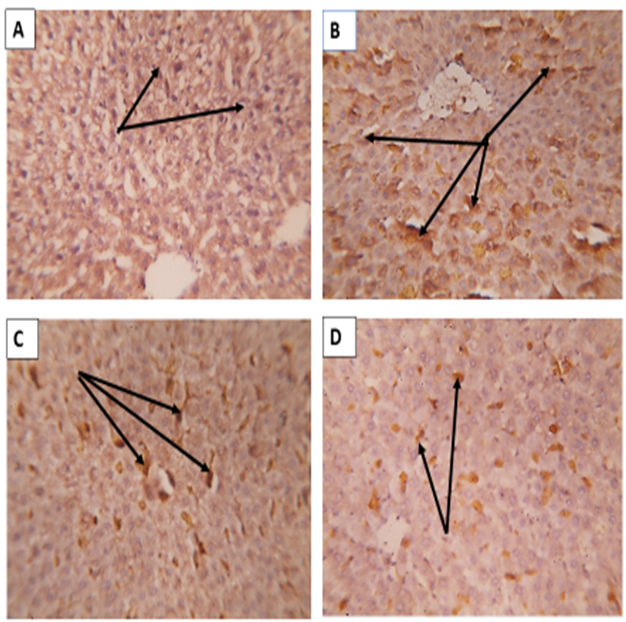
| Figure 9: : Sections in the liver of rats immunostained with CD 68 ( phagocytic Kupffer cells) showing ;(A)Healthy G0 Control group showed negative or few CD68 positive cells ( arrows). (B) Iron intoxicated group: showing numerous large sized CD68 immuno-positive cells ( arrows).(C) NAC treated group (G2): showed less smaller sized Cd68 positive cells. (D)Acetylated whey treated group (G3): showing evident decrease in size and number of Cd68 positive cells. (CD68 immunostaining X400) |
 |
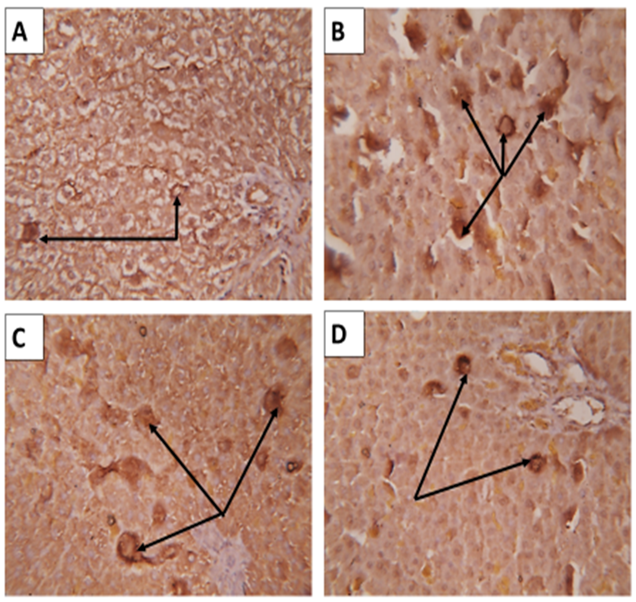
| Figure 10: Sections in rat liver immunostained by CD163; A) Healthy G0 control group showed merely negative reaction. B) Iron over load intoxicated group G1 showing intense positive reaction (arrows). C) NAC treated G2 group showed a decrease in the positive reaction than the only iron intoxicated G1 (arrows). D) ABW treated group G3 showing a decrease in the intensity of the positive distribution reaction than the previous G2 treated group (arrows). (X400) |
 |
| Figure 11: Sections from rat liver immuno-stained for Caspase- 3 immuno-expressions showing ; (A) Healthy control group (G0) showing faint defuse staining in some hepatocytes ( arrows). (B) Iron over load intoxicated (G1) group showing an increase in hepatocytes with intensive positive caspase-3 immuno-expression.( arrows). (C) The NAC treated intoxicated (G2) group showing mild decrease in the intensity of positively stained hepatocytes (arrows) than that of G1 group. (D) The acetylated whey (ABW) treated during iron intoxicated group (G3) showed marked decrease in the intensity and number of the positively stained hepatocytes (arrows) than those of the previous G2 treated group. (X400) |
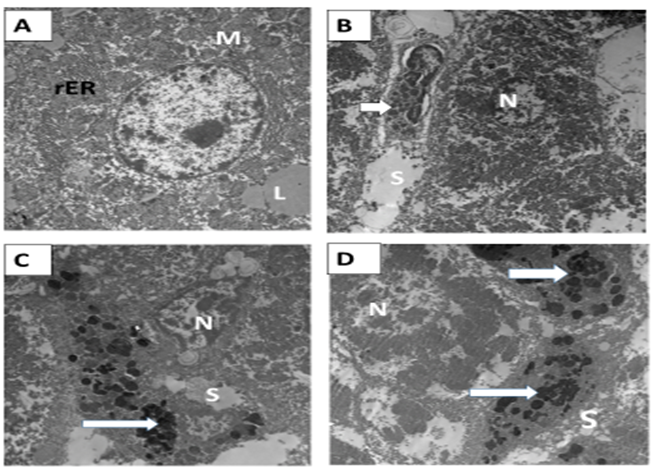 |
| Figure 12: Electron micrograph in rat liver A&B) Healthy control G0 group showing; normal hepatocyte with euchromatic nucleus (N). The cytoplasm contained well developed RER (rER) and mitochondria (M). Lipid droplets (L) Von Kupffer cell (arrow) were seen lining the blood sinusoids (S). C&D) Ferrous intoxicated G1 group showing; the nucleus (N) of hepatocyte appeared irregular, heterochromatic, and surrounded by many vacuoles. Numerous mitochondria (M) were observed amalgamated with each other with ill-defined cristae and electron-dense matrix. The Von Kupffer cell was engorged with numerous and irregular secondary lysosomes (arrows). (X4800, 3600 ,4800 and 3600) . |
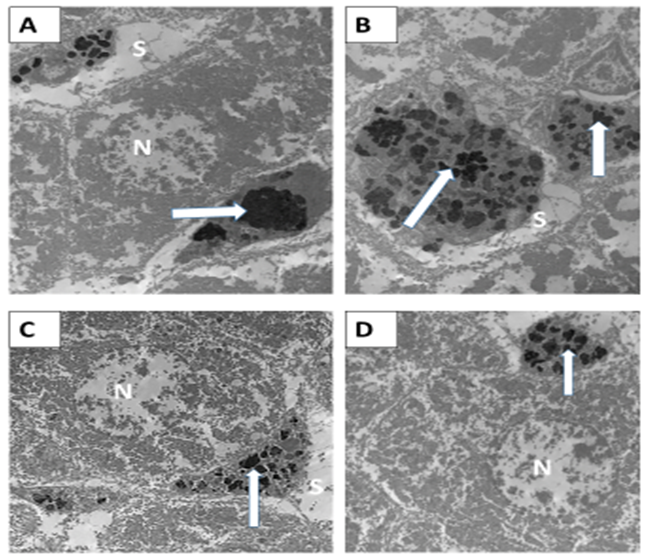 |
| Figure 13: (A&B) N-acetyl Cysteine Iron treated group (G2) showing; the Von Kupffer cells appeared still engorged with secondary lysosomes (arrows) similar to some extent to the iron intoxicated group. The mitochondria (M) still appeared smaller in size and amalgamated with each other. (C&D) Acetylated peptide and Iron treated group (G3) showing; the Von Kupffer cells (arrows) appeared smaller in size and not engorged with secondary lysosomes compared to the previous treated groups. The mitochondria(M) appeared separated from each other with ill-defined cristae. (X3600). |




































