Open Access
Research Article
Max Screen
ISSN: 2394-6512
Copyright: © 2023 Andrea Celeste Isaguirre. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Candida albicans is an opportunistic agent that can produce systemic infections in immunosuppressed patients. By varying its cell wall antigens this fungus can evade the immunological response by varying its cell wall antigens. On the other hand, Larrea divaricata Cav. (Jarilla) is a widely spread plant in America and it is used in folk medicine to treat several pathologies. It has been shown that antibodies against Jarilla proteins of crude extract (JPCE) cross-react with proteins of other microorganisms such as Gram-negative bacteria.
Herein we assessed the contribution of anti-JPCE antibodies in the opsonophagocytosis of C. albicans by murine macrophages, and in the recognition of Candida virulence factors. The reactivity of anti-JPCE IgG antibodies against cell and wall proteins suggest that these proteins induce a response that could enhance opsonic recognition of this yeast, which is important for its elimination. Opsonophagocytosis assays also show that these antibodies could increase yeast intake. These heterologous antibodies also neutralize the effects of somatic total proteins of C. albicans that induce death of the macrophage by the fungus. L. divaricata proteins could act by enhancing the production of heterologous protective and/or neutralizing antibodies to Candida albicans.
Keywords: Candida Albicans; Larrea divaricata Cav.; Cross-Reaction; Immunogenic Proteins; Opsonophagocytosis
Fungi cause approximately more than one million lethal infections each year. Most serious fungal infections are caused by Candida spp. Among these species, C. albicans is the most commonly isolated and represents the most common fungal opportunistic pathogen that causes systemic infections in immunosuppressed patients. The high mortality rates associated with invasive fungal diseases and septicemia reflect the urgent need for protective vaccines [1-3]
Candida albicans is a polymorphic fungus that can grow either as ovoid-shaped budding yeast, as elongated ellipsoid cells with constrictions at the septa (pseudohyphae), or as parallel-walled true hyphae. Further morphologies include chlamydospores, which are thick-walled spore-like structures. While yeast and true hyphae are regularly observed during infection, the role of pseudohyphae is rather unclear and chlamydospores have not been observed in patient samples. The transition between yeast and hyphal growth forms is termed dimorphism. The hyphal form is more invasive than the yeast form, but both forms are important in pathogenesis of candidiasis. On the other hand, the smaller yeast form is believed to represent the primarily form involved in dissemination of candidiasis [4].
Candida albicans is surrounded by a very resistant cell wall responsible for interactions with the host, including triggering and modulating its immune response. This seems to be based on a complex interaction between natural and adaptive immune response [5]. Indeed, the main immune defense mechanism against C. albicans is based on cellular response [6].
The importance of neutrophils in phagocytosis of C. albicans is evident from the increased rate of infections seen in patients with severe neutropenia. The major response associated with phagocytosis of microbial pathogens is the oxygen-dependent respiratory burst with the generation of cytotoxic reactive oxygen species (ROS) [7]. Cells of the phagocytic system can recognize C. albicans through multiple classes of receptors [8]. While these receptors can induce phagocytosis, the efficiency of uptake in both neutrophils and macrophages can significantly increase when C. albicans is opsonized by antibodies [9] or complement proteins [10]. Moreover, several therapeutic monoclonal antibodies exert their protective effects by opsonizing cells for phagocytosis [11]. It has been demonstrated that neutrophils present a significant increase in the level of phagocytosis of antibody-opsonized microorganisms [12].
Larrea divaricata Cav. (commonly known as “Jarilla”) is a plant from the Zygophyllaceae family that is widely distributed in South America. In particular, in Argentina, from Salta to Chubut, between 500 to 2300 m of altitude [13]. Extracts from L. divaricata Cav. have anti-tumor activities against different cell lines [14], and also have anti-fungal and anti-bacterial properties [15]. Our research team has shown the in vitro and in vivo effect of jarilla aqueous extracts on the immune system. Furthermore, in vivo activation of macrophages against C. albicans has also been informed [16]. Jarilla extract caused macrophage priming, increased phagocytosis and production of reactive oxygen species (ROS) and nitric oxide (.NO), and did not kill the macrophage [17,18].
Additionally, proteins isolated from the crude extracts of jarilla have been used for the immunogenic characterization of Jarilla and their relationship with antigens from different pathogens such as Pseudomonas aeruginosa, Proteus vulgaris, Klebsiella pneumonia and Escherichia coli O157 H:7 [19,20]. Interestingly, recent studies have shown that sera obtained from mouse inoculated with partially purified JPCE, can inhibit the cytotoxic and hemolytic activity caused by cellular and extracellular proteins obtained from P. aeruginosa [21]. These sera also contain opsonizing antibodies that stimulate the phagocytic activity of murine macrophages against this bacterium [22]. However, it remains to be determined whether antibodies generated against JPCE can enhance the opsonization and phagocytosis of yeasts by myeloid cells such as mouse peritoneal macrophages
Herein we aimed at identifying immunogenic proteins of JPCE, and the determination of their antigenic similarity with total-cell proteins (TCP) and surface-cell proteins (SCP) of C. albicans. This is critical for the understanding about how the immune response against JPCE can protect against infections by Candida albicans.
The aerial parts of L. divaricata Cav. were collected from Nogolí, San Luis (32° 55´ 15,7´´ S; 66° 19´ 18,6´´ W; altitude 799 m a.s.l).The crude extract was obtained and the proteins were partially purified, but only with 10 kDa cut-off membrane concentrators. The protein concentrations of JPCE was determined by the Bradford method using an albumin standard of 1.0 mg ml-1 .
The reference strain C. albicans ATCC 36801 (type strain) obtained from the American Type Culture Collection (Maryland USA) was kindly provided by Dr. Claudia Sotomayor (CIBICI, Center for Research in Clinical Biochemistry and Immunology, National University of Córdoba, Argentina).
The strain was grown on Sabouraud Glucose agar medium at 37°C for 24 h. For the tests, inocula were used in an exponential phase in order to obtain a predominance of yeasts with blastoconidia morphology. The morphological characteristic was assessed by optical microscopy. The cells were used to obtain total and surface proteins.
Total cell proteins, TCP: The yeasts grown on Sabouraud Glucose agar were harvested and washed 3 times with sterile PBS by successive centrifugations at 7000 x g for 5 min at 4 °C. Then, the cell pellet was suspended in 15 ml of sonication buffer and sonicated at 80 kHz for 10 min (50% cycles).and sonicated at 80 kHz for 10 min (50% cycles). The sonicate was centrifuged at low speed (20 min at 1000 x g) to remove large fragments and whole cells. The supernatant was used as antigen preparation and stored at -20 °C until use.
Surface cell proteins, SCP: The yeasts grown on Sabouraud Glucose agar were harvested and washed 3 times with sterile PBS. The cell pellet was suspended in 15 ml of sonication buffer and sonicated at 80 kHz for 10 min (50% cycles). The sonicate was centrifuged at low speed. Cell wall proteins were extracted of cell pellet, for 10 min at 100°C with lysis buffer (50 mmol l-1 Tris-HCl, pH 8.0, 0.1 mol l-1 EDTA, 2% SDS, 10 mmol l-1 2-mercaptoethanol) [23]. The proteins were obtained by centrifugation at 1650 x g for 5 min. The supernatant was stored at -20°C until use. The protein concentrations of TCP and SCP proteins were determined by the Bradford method using an albumin standard of 1.0 mg ml-1 .
Batches of male Balb/c mice of 18 to 20 g of body weight were housed and cared at the Animal Resource Facilities, Faculty of Chemistry, Biochemistry, and Pharmacy, National University of San Luis (protocol approval code B115/12, Institutional Committee for the Use and Care of Animals, UNSL).
The protein analysis and the immunization protocol were carried out according to Canale et al. [22]. The JPCE, TCP, and SCP protein samples were subjected to 12% polyacrylamide gel electrophoresis using the discontinuous Laemmli system.
On the other hand, groups of six mice were immunized subcutaneously at a 21-day interval, with two doses of 0.2 ml of JPCE, TCP and SCP (3 mg ml-1) in adjuvant AlPO4 (1:1) [22].
ELISA plates (Costar, Cambridge, MA, USA) were sensitized overnight at 4 °C with 50 μg well-1 of JPCE in carbonate/ bicarbonate buffer pH 9.6. The plates were rinsed with PBS-Tween 0.05% and blocked with PBS pH 7.2-1% skim milk for 1 h at 37 °C. The blocking solution was removed, and the plates were rinsed again.
The inhibition experiments were carried out as previously described by Mattar de Anaya et al. [19], pre-incubating equal volumes of different concentrations of JPCE, TCP or SCP (10, 5, 2.5 μg ml-1) and diluted anti-JPCE serum (titer 1/800) in PBS, for 1 h at 37°C. The dilution of this antiserum was previously determined by titration assays. After incubation, the samples were centrifuged at 18500 x g and 100 µl of each supernatant were transferred to the wells sensitized with JPCE. The non-inhibited anti-JPCE was used as a positive control. After incubation for 24 h at 4 °C, the plates were rinsed 5 times with PBS-Tween. A goat anti-mouse IgG, Horseradish Peroxidase (HRP) conjugate at 1:300 dilution was added and incubated at 37 °C for 1 h. Detection with HRP was performed, according to Canale et al. [22].
ELISA plates were sensitized with 50 μg well-1 of JPCE or SCP in carbonate/bicarbonate buffer pH 9.6, overnight at 4 °C. The plates were rinsed with PBS-Tween 0.05% and blocked with PBS pH 7.2 containing 1% skim milk for 1 h at 37 °C. After the removal of the blocking solution, the plates were rinsed 3 times with PBS-Tween. For the qualitative analysis, sera from mice immunized with SCP or JPCE, were diluted 1:50 in PBS were tested. For the semiquantitative analysis, serial dilutions 1:2 in PBS pH 7.2 from a dilution starting at 1:50 were performed. Then the technique described in previous section was followed.
The optical density was compared with that obtained with pre-immune sera (PI).
The antigens from SCP were identified by specific anti-JPCE antibodies obtained from animals immunized with JPCE using the Western blot technique according to Canale et al. [22].
The possible opsonizing effect of anti-JPCE sera on phagocytosis of C. albicans by mouse peritoneal macrophages was examined by live-cell imaging using mouse macrophages
Peritoneal macrophages (0.5 ml, 1×106 cells ml-1) were placed on microscope slides and incubated for 1 h at 37 °C. The slides were rinsed with HBSS (Hank's Balanced Salt Solution, Sigma Aldrich, St. Louis, MO, USA) to eliminate those cells that did not adhere and subsequently, incubated with the opsonize C. albicans suspension with different sera (anti-JPCE, anti-C. albicans and PI sera) at multiplicities of infection (MOI) of 1:10 . Opsonized yeasts with PI sera were used as control. After 1 h of incubation, the slides were rinsed with cold PBS. The slides were dried, fixed with methanol, and stained with 3% Giemsa in PBS. The location of the yeasts was observed by optical microscopy. The results were expressed as normalized values with respect to those obtained in opsonophagocytosis test with PI sera.
The ability of purified macrophages to phagocyte C. albicans was evaluated by flow cytometry. Nine million yeasts were rinsed and resuspended in 1 ml of PBS. Yeasts were fluorescently labeled with 1.5 μl of succinimidylcarboxyfluoresceindiacetate (CFSE), vortexed, incubated for 20 min at 37 °C, and opsonized with anti-JPCE serum. Subsequently, 1x106 macrophages were incubated with opsonized yeast (MOI 1:3) in 24 wells plates for 40 min. After that time, cells were placed on ice and harvested by scraping. To quench the fluorescence of the non-ingested yeasts, cells were incubated with 1 ml of trypan blue (500 μg ml-1) for 5 min at 25 °C. Then the macrophages were rinsed twice with cold PBS and resuspended in 300 μl of FACS buffer. The incorporation of fluorescence by macrophages was analyzed by flow cytometry (BD FACSCalibur).
Groups of 4 animals were injected intraperitoneally with 1 ml of 10% peptone protease (Difco) in sterile PBS. After 48 h, the peritoneal cells were collected by washing the peritoneal cavity with sterile HBSS. The cells were rinsed twice and re-suspended in DMEM (Dulbecco's Modified Eagle Medium) supplemented with glutamine, fetal bovine serum, and a cocktail of antibiotics, and adjusted to 1x106 cells ml-1. The macrophages were purified from peritoneal cells by adhesion onto the bottom of 96-well culture plates for 2 h at 37 °C. The non-adherent cells were removed by rinsing the plates with Hank's solution.
The reduction of bromide of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was tested as previously described by Martino et al. (Martino et al. 2012). The macrophages were treated with TCP or SCP (300, 190, 95, 45 and 25 μg ml-1) and incubated with 100 μl of DMEM and 10 μl of 5 mg ml-1 of MTT for 4 h. The formed purple formazan was solubilized by adding isopropanol-acid (50 ml 2 mol l-1HCl in 2.5 l Isopropanol). The absorbance was measured using a plate reader (Microplate Reader Benchmark, Bio-Rab) at a wavelength λ of 550nm. The untreated cells were used as a control of viability (100%) and the results are shown as relative viability percentage with respect to the control.
For this assay, cell purification by adhesion was performed in hemolysis tubes. After purification, the cells (1x106 macrophages percondition) were incubated for 1 h with different concentrations of TCP (300, 190, 95, 45 and 25 μg ml-1) at 37 °C. Subsequently, the cells were washed and incubated with 200 μl of 0.1% NBT (Nitro Blue Tetrazolium) and 100 μl of zymosan (Zim) (1 mg ml-1) for 1 h at 37 °C. Then the cells were lysed with 500 μl of 1 mol l-1 HCl and centrifuged at 5000 x g for 5 min. The supernatant was discarded and 300 μl of dioxane was added to solubilize the crystals of reduced NBT (formazan). Finally, 200 μl of solubilized formazan was transferred to a 96 well plate and the absorbance was measured at a wavelength of 550 nm. The results are expressed in nM of formazan produced per every million cells.
Two million cells per condition were purified by 96-well plate adhesion. The cells were incubated with 100 μl of different concentrations of TCP (300, 190, 95, 45 and 25 μg ml-1) and 100 μl of LPS (10 ng ml-1) to stimulate the macrophage activation for 24 hours at 37 °C. Then, 100 μl of the supernatant were taken and plated in 96 wells, to which 100 μl of the Griess reagent was added Reagent A: sulfanilamide (2% w/v in 5% HCl) + Reagent B: N-1- (naphthyl) ethylenediaminedihydrochloride (0.1% w/v in 5% HCl). The mixture was incubated for 10 min in the dark, at room temperature for the subsequent reading of the absorbance at a λ = 550 nm. The results are shown as a percentage of nitrite production compared to the control.
To evaluate the neutralization of the effects of TCP on macrophages, the antigenic preparations were incubated with a dilution of 1/50 mouse anti-JPCE sera for 1 h at 37 °C. After incubation, the samples were centrifuged at 16000 x g and the supernatant obtained was tested to demonstrate the neutralization utilizing specific antibodies and evidenced by the decrease or the disappearance of the observed effect.
The similarity of protein bands of SDS-PAGE was expressed utilizing the Dice Similarity Coefficient (DSC): 2nAB/(nA+nB) *100,where nAB is the number of common bands, and nA and nB are the numbers of total bands in profiles A and B.
In the experiments, multiple pools of PI, anti-JPCE, anti-SCP and anti-TCP sera were used. Also, at least two different populations of macrophages were used in each test. Differences between group means were assessed using one-way ANOVA followed by multiple comparisons by Tukey test.
In the case of comparing only two groups, unpaired Student t test was used for non-homogeneous variances. The outliers were identified using the statistical method of Grubb. All statistical analyses were performed with Graph Pad Prism version 9.00 for Windows (Graph Pad Software, San Diego California USA). A value p≤0.05 was considered statistically significant. *, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001. The expressed results represent the mean value of replicates or the most representative experience.
Protein profiles were analyzed by SDS-PAGE. This method allowed the identification of protein bands whose apparent molecular masses ranged between 150 and 20 kDa. A large number of protein´s bands were identified in the different antigenic preparations: JPCE, 15 bands; TCP, 35 bands, and SCP, 32 bands (Figure 1). In the case of JPCE, bands with apparent molecular mass (Mm) between 150 and 20 kDa were identified.
After comparing these electrophoretic profiles, common bands were identified between the cellular proteins of C. albicans and JPCE. JPCE proteins have a percentage of similarity of 48% and 42 % with TCP and SCP, respectively. The common bands for SCP-JPCE were those of apparent Mm of 113, 109, 91, 75, 70, 62, 58, 34, 22, 20 kDa and for TCP, in addition to those named above, bands 37 and 31 kDa, were observed.
ELISA inhibition tests were performed to establish the cross-reactivity between jarilla and C. albicans proteins using a polyclonal antiserum anti-JPCE. The percentage of inhibition was greater than 50% when the anti-JPCE sera were neutralized with JPCE (66%, 63%, and 58%) and SCP (65%, 56%, and 54%). TCP caused a 53% inhibition when used at 2.5 µg ml-1, whereas when used at 10 and 5 µg ml-1 it only caused an inhibition of 33% and 31%, respectively
The immunogenicity of the JPCE and SCP antigen preparations was studied using a qualitative ELISA test. The optical density was compared with that obtained with pre-immune sera (PI).Results are shown in Figure 2 (A y B).
Western-Blot assays were performed to identify the immunoreactive SCP of C. albicans with sera from animals immunized with JPCE (title of 1/1600). The results obtained are shown in Figure 3.These assays showed thirteen immunoreactive bands (97, 90, 80, 65, 55, 51, 48, 38, 35, 30, 22, 20 and 18kDa).
Taking into account that the sera from animals immunized with JPCE presented cross-reactivity with SCP, the opsonization capacity was assessed. The opsonophagocytosis test of C. albicans was carried out with the different sera (anti-JPCE and anti-SCP). The mouse peritoneal macrophages (Mϕ) treated with C. albicans opsonized with anti-SCP sera showed 11% polyphagocytosis (many phagocytosed cells), 22% associated cells, 18% phagocytosis and 30% necrosis. When the yeasts were opsonized with anti-JPCE sera, the results were 17% polyphagocytosis, 25% associated cells, 20% phagocytosis and 23% necrosis. The results observed in the control were 6, 17, 10 and 19%, for polyphagocytosis, associated cells, phagocytosis and necrosis respectively. Table 1 shows the normalized data compared to the data obtained with the PI sera. The table shows a high polyphagocytosis when C. albicans was opsonized with anti-JPCE (2.8 times) versus data obtained with anti-SCP (1.8 times). In associated cells the results were of 1.5 and 1.3 between anti-JPCE and anti-SCP, respectively. In phagocytosis, the results were of 1.8 and 1.9 between anti-JPCE and anti-SCP, respectively. In all cases the necrotic effect of C. albicans on the Mϕ was observed. It should be noted that anti-JPCE antibodies decreased the effect produced by anti-SCP by 0.4 times.
Our optical microscopy data showed that Jarilla anti-protein antibodies recognize proteins in C. albicans, thus enhancing the yeast phagocytosis. Figure 4 (A-D) shows representative images of Mϕ´s morphologies as observed in the opsonophagocytic assay
To confirm the opsonophagocytosis of C. albicans by the anti-JPCE sera, flow cytometry assay was performed. The results are shown in Figure 5. The test showed that the anti-JPCE serum increased the phagocytosis of the yeasts by 12% compared to the anti-SCP serum.
To evaluate the cytotoxic effect of C. albicans proteins the metabolic reduction assay of MTT was carried out in mouse Mϕ, treated with different concentrations of TCP and SCP.
A dose-dependent cytotoxicity effect was observed with TCP. TCP caused a dose-dependent reduction on Mϕ viability: 40% at 300 μg ml-1, 62% at 190 μg ml-1, 64% at 95 μg ml-1, 73% at 45 μg ml-1, and 81% at 25 μg ml-1, and this effect was no observed with SCP (Figure 6.). Because the cytotoxic effect was more evident and specific for the total and non-surface cellular proteins, the neutralization of TCP was then tested.
Macrophages were treated with 95 μg ml-1 of TCP (62% of cell viability). The proteins were incubated with anti-JPCE and anti-TCP sera (1/50). Anti-jarilla and anti-TCP sera significantly increased the percentage of cell viability up to 84 and 82%, respectively (p=0.009). Figure 7 shows the values of viability obtained with TCP and the effect of neutralization by anti-TCP and anti-JPCE sera.
To evaluate the effect of C. albicans cellular proteins on the production of superoxide anion and H2O2 by the Mϕ, the reduction of NBT was determined. The tested concentrations of TCP decreased significantly (p≤ 0.02) the phagocytic activity compared to the control, but there was no significant difference between the different concentrations tested.
The nmoles of reduced NBT, obtained with Mϕ treated with 95 μgml-1 TCP previously incubated with anti-JPCE (1/50) sera did not neutralize the effect with respect to the control group (Mϕ with TCP without anti-JPCE).
The nitrite production, as an indirect test of NO production, by macrophages treated with different concentrations of TCP, was significantly reduced by 45 and 25 μg ml-1 concentrations. The neutralizing effect of the anti-JPCE (1/50) sera was not observed when they were pre-incubated with TCP (data not shown).
Our research team has demonstrated cross-reactivity between proteins of JPCE and cellular antigens and extracellular products of P. aeruginosa, respectively [19,24]. However, there are no previous studies on the immunogenic relationship between Jarilla proteins and C. albicans.
From the analysis of protein profiles by SDS-PAGE of JPCE, bands with apparent molecular mass (Mm) between 150 and 20 kDa were identified, and these results are consistent with those reported by Mattar de Anaya et al. [19]. In this sense, the bands that Jarilla shares with SCP and TCP, whose Mm is between 80 and 15 kDa, would correspond to adhesins and receptors [6].
The study of the immunogenic capacity of JPCE proteins by ELISA, showed that the molecular mimicry, at the concentrations tested, was greater with SCP than with TCP of C. albicans.
When JPCE was used as a sensitizing antigen, we observed that the IgG titer of the sera obtained from mice immunized with JPCE (anti-JPCE) and SCP (anti-SCP) were higher than that of the PI serum (p< 0.0001 for both immunological reactions). When SCP was used as a sensitizing antigen, the responses obtained were significantly higher with respect to the control (p< 0.001 for both immunological reactions). However, when homologous and heterologous immunogenic responses against JPCE and SCP antigens were compared, no significant difference was observed. Moreover, there was no significant difference between the average titers obtained in anti-JPCE sera with both sensitizing antigens, JPCE (1/1600) and SCP (1/1600).
Considering the greater protein profile similarity with SCP, the immunological cross-reactivity between anti-JPCE with SCP was evaluated. The values obtained were comparable to the homologous reaction. This data supports our previous report showing the high immunogenicity of Jarilla´s proteins in mice [24]; and the immunological heterologous reactivity with SCP of C. albicans.
Taken together these results indicate the recognition of SCP of C. albicans by heterologous antibodies against jarilla and the probable presence, therefore, of common antigens between these two eukaryotic organisms studied. It is known that the molecular machinery of many cellular processes is conserved in bacteria, yeasts, plants and also mammals. This is illustrated by the fact that higher eukaryotic genes have been introduced in yeasts for systematic analysis of their function [3]. This could be a probable explanation of the presence of common antigens. It is well known that the surface glycoproteins of unicellular fungi play an important role in the host-pathogen interaction. The main glycoproteins (mannan oligosaccharide polymers covalently linked to proteins) are responsible for the immunogenicity of the cell walls [6].
Western-Blot assays showed thirteen immunoreactive bands. It should be noted that the 90 kDa band possibly corresponds to Hsp90, which is the thermal shock protein necessary for yeast. The Hsp90 protein plays an essential role in the resistance of biofilms of C. albicans to certain antifungal agents. Blocking of this protein decreases the production of glucans and therefore decreases the amount and density of the extracellular matrix, thus facilitating the input of therapeutic agents [25]. In addition, this band has been characterized in both yeast and hypha [26].
An 81 kDa monomeric protein with hydrolase activity would be associated with host-cell membrane damage, adhesion and penetration, thus being an important virulence factor of C. albicans [6].
The band 65 kDa could correspond to a mannoprotein of the cell surface of C. albicans that plays a major role in the host-parasite interaction and has been characterized as a major target of the human immune response against C. albicans [27].
El Moudni et al. [28] showed that C. albicans cells contain high levels of metallopeptidase. This enzyme displays an electrophoretic mobility equivalent to about 52 kDa. Identification of such a type of enzymatic activity could have important implications for the ability of C. albicans to invade host tissues by degrading the extracellular matrix [6].
Other research teams have identified a protein with high immunogenicity whose molecular mass was 50 kDa. This protein is only present in the yeast state [26].
The protein bands between 38 and 49 kDa could correspond to secreted aspartic proteases (Sap) isoforms 1-3, present in yeast. Sap are enzymes involved in tissue invasion and also in adhesion to epithelial cells, among other functions [29,30].
On the other hand, the 38 kDa band could correspond to a protein from the lipase family [6]. The anti-JPCE-immunoreactive protein bands observed at 14 and 18 kDa could correspond to the peptides with the highest immunogenic capacity. These are products of denaturation of greater molecular mass proteins.
These results led to the proteomic study of Jarilla´s immunoreactive bands, which are cross-reactive with C. albican´s protein virulence factors.
Opsonization assays were tested through optical microscopy. Our optical microscopy data showed that Jarilla anti-protein antibodies recognize proteins in C. albicans, thus enhancing the yeast phagocytosis.
Macrophage necrosis process is an immune evasion mechanism due to the intake of the opsonized fungus. This process was decreased by anti-jarilla antibodies, suggesting a cross-reaction with proteins of C. albicans that promote the mechanism of macrophage cellular death. Among these proteins, proteases (Saps) [30,32] have been suggested that can act as cytolysins in macrophages after phagocytosis of Candida [32].
These results, confirmed by flow cytometry, indicate that anti-JPCE antibodies that recognize SCP could have opsonophagocytic action on C. albicans. This characteristic may provide a protective mechanism against yeast in the early stages of infection [11]. Rudkin and colleagues demonstrated C. albicans yeast and hyphal cells treated with anti-whole cell mAbs or anti-Hyr1 mAbs were phagocytosed more quickly compared to non-opsonized cells [3]. The opsonophagocytic activity was also suggested by the protective effect of selected antibodies in a murine model with disseminated candidiasis [33].
In order to specify the antigenic similarity between jarilla proteins and C. albicans in yeast state, with respect to those involved in the viability and production of reactive oxygen and nitrogen species, neutralization experiments were carried out in macrophages. Total cellular proteins of C. albicans decreased macrophage viability indicating the presence of proteins causing cell death. Peroumal et al. showed that the inhibition of specific proteins with pepstatin prevented the early processes of apoptosis and necrosis in mouse peritoneal macrophages infected with C. albicans [34]. Because TCP showed a significant alteration of mitochondrial functionality (MTT reduction) with respect to that produced by SCP under the conditions tested, TCP neutralization tests were performed. The results showed that neutralization of these proteins with the anti-JPCE sera was similar to that produced by the homologous reaction. Therefore, the jarilla crude extract might have antigenic determinants common to those found in C. albicans proteins that participate in immune evasion mechanisms [32].
Candida albicans proteins at the concentrations tested, reduced phagocytic activity and NO production. These effects of C. albicans proteins may favor the survival of yeast within the macrophage during the phagocytosis process. Anti-JPCE could not significantly inhibit this effect of C. albicans. This could be due to an absence in the antigenic similarity with the C. albicans proteins involved in such mechanisms, or due to the minimal dilution used for the tested sera.
There is an increasing awareness of the concept, as well as experimental documentation of, heterologous immunity and cross-reactivity of adaptive immune lymphocytes in protection against infection [35].
Therefore, a very interesting proposal would be to study these cross-reactivities between vegetable proteins, as harmless and highly specific factors, that can redound in immunoprotective effects and in the resistance against C. albicans infections.
All authors of this manuscript have the right to authorship and have each approved the final version of the submitted manuscript.
All authors warranty that they have the authority to publish the material.
All authors warranty that the paper, or one substantially the same, has neither been published previously, nor is being considered for publication elsewhere.
The authors would like to acknowledge the financial support received from the project of CyT PROICO 02-3018 from the National University of San Luis (FQByF), Argentina.
No conflict of interest to be declared.

![]()

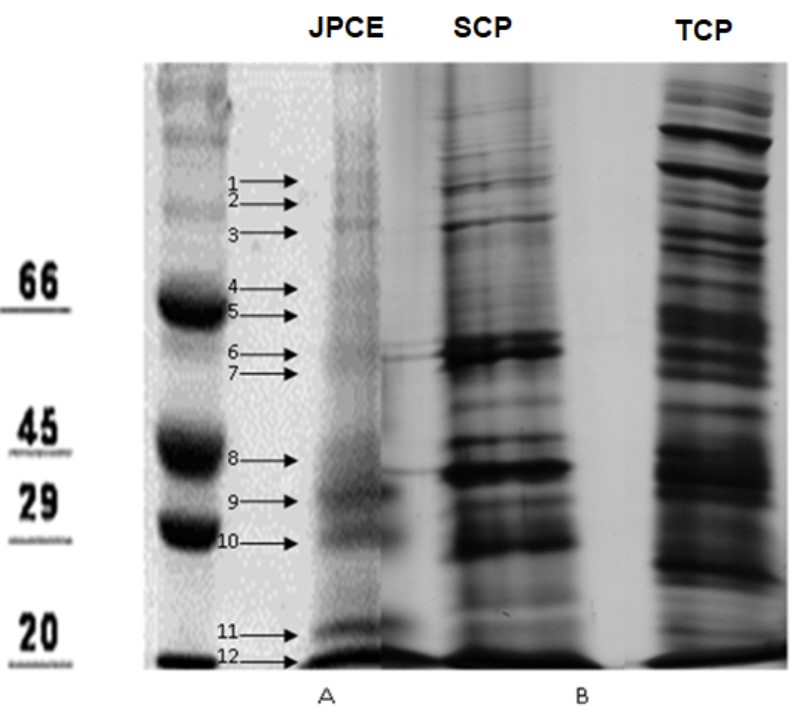 |
| Figure 1: SDS- Protein profiles of L. divaricata and C. albicans. JPCE, Jarilla proteins of crude extract; SCP, superficial cell proteins of C. albicans; TCP, total cell proteins of C. albicans. The arrows indicate the common protein bands between JPCE, SCP, and TCP. Molecular weight markers expressed in kilodalton (kDa) |
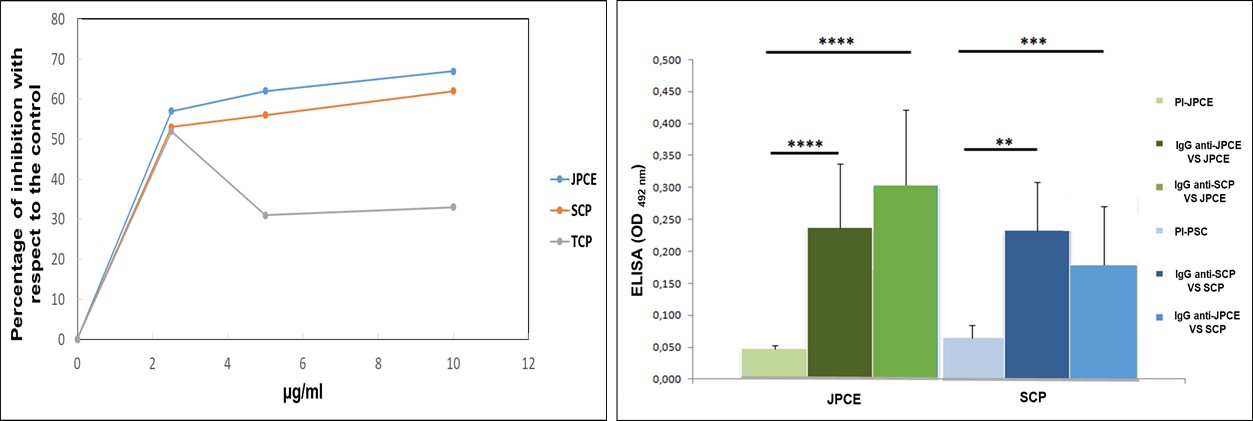 |
| Figure 2: A- ELISA inhibition test. Antigenic preparations: Jarilla proteins of crude extract (JPCE), surface cell protein (SCP) and ,total cell proteins (TCP) in three different concentrations (10, 5, 2.5 μg ml-1) were incubated with anti-JPCE serum at 1/800. This dilution of this antiserum was previously determined by titration assays. B- Qualitative ELISA test. Levels of anti-JPCE IgG and anti-SCP IgG using JPCE and SCP as coating antigens, respectively. Results are shown as the mean values of absorbance at 492 nm. ****, p< 0.0001 and ***, p< 0.001 |
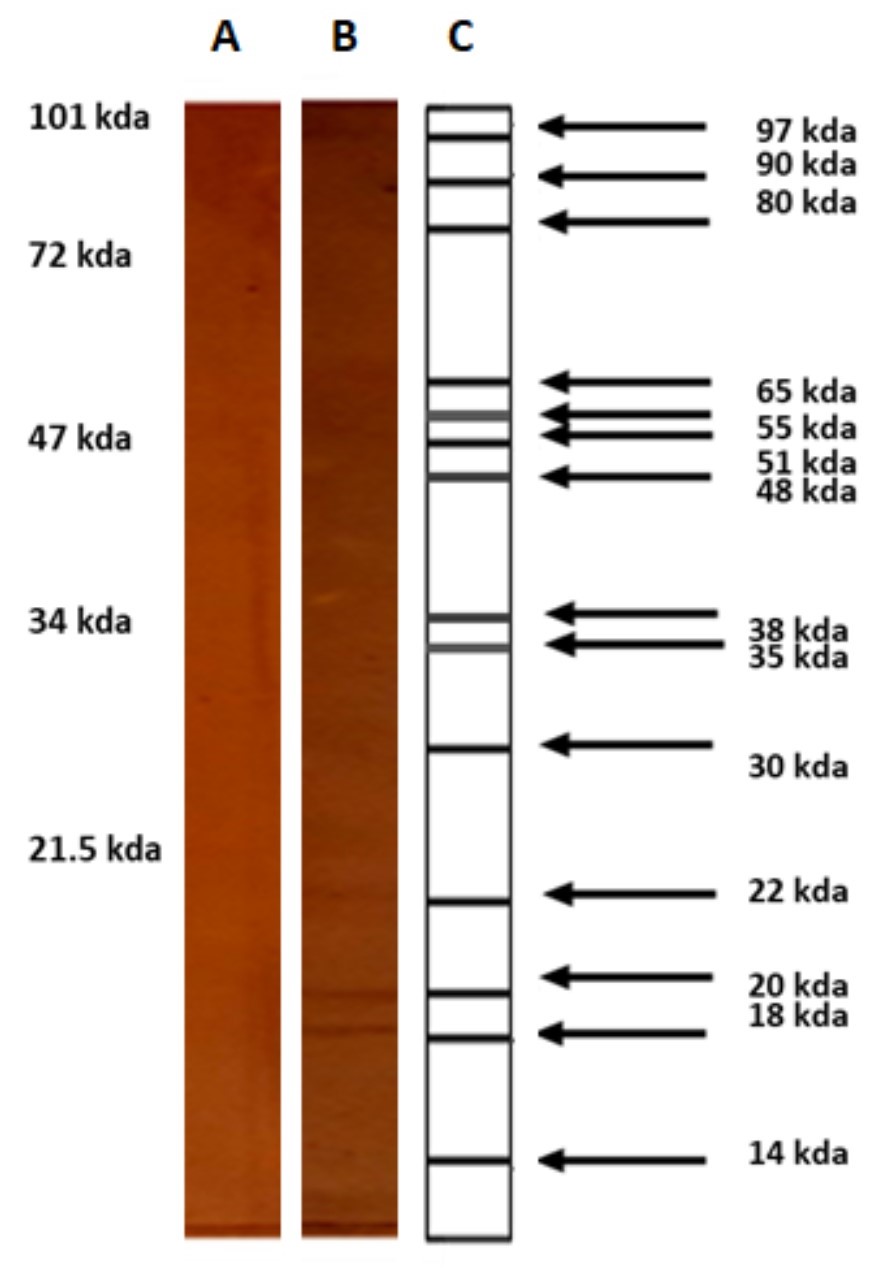 |
| Figure 3: Western blot. Analysis of immunoreactivity of surface cell proteins (SCP) of C. albicans with pre-immune sera (negative control)(A) and anti-JPCE sera obtained of mice immunized with Jarilla proteins of crude extract (B). C, molecular weight of the bands recognized by anti-JPCE sera and on the left, the molecular weight markers, expressed in kilodaltons (kDa) |
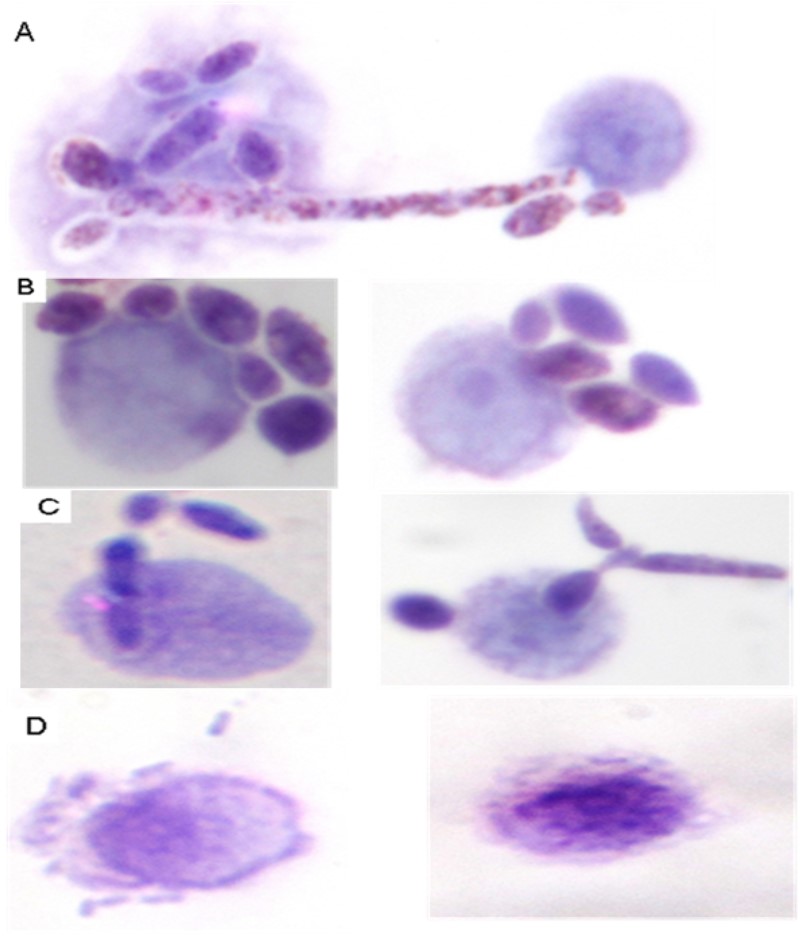 |
| Figure 4: Optical microscopy. Cross-reaction of anti-JPCE with C. albicans. On slide-laid, murine peritoneal macrophages were incubated with a suspension of yeast cells opsonized with different sera (anti-JPCE, anti-C. albicans and pre-immune sera (PI)) at multiplicities of infection (MOI) from 1:10. As a control, yeasts opsonized with PI sera were used. The images show: A, polyphagocytosis; B, associated cells; C, phagocytosis; D, necrosis |
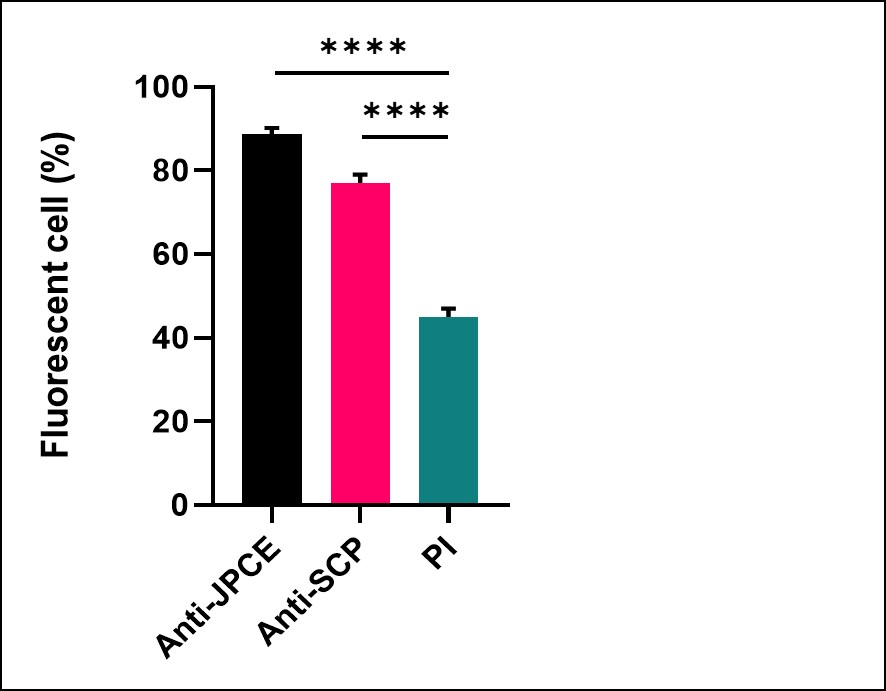 |
| Figure 5: Opsonophagocytosis assay by flow cytometry. 1x107 cells of C. albicans were fluorescently labeled with 1.5 μl CSFE and opsonized with sera (anti-JPCE or anti-Surface Cell Protein (SCP) of C. albicans) from a pool of six mice immunized with Jarilla proteins of crude extract or SCP. They were incubated with mouse peritoneal macrophages (M?) purified at multiplicities of infection (MOI) 1:3 for 40 minutes at 37o C to allow phagocytosis. The graph shows the percentage of yeasts evaluated that were positive for fluorescence under different opsonization conditions. PI: pre- immune sera****, p < 0.0001 |
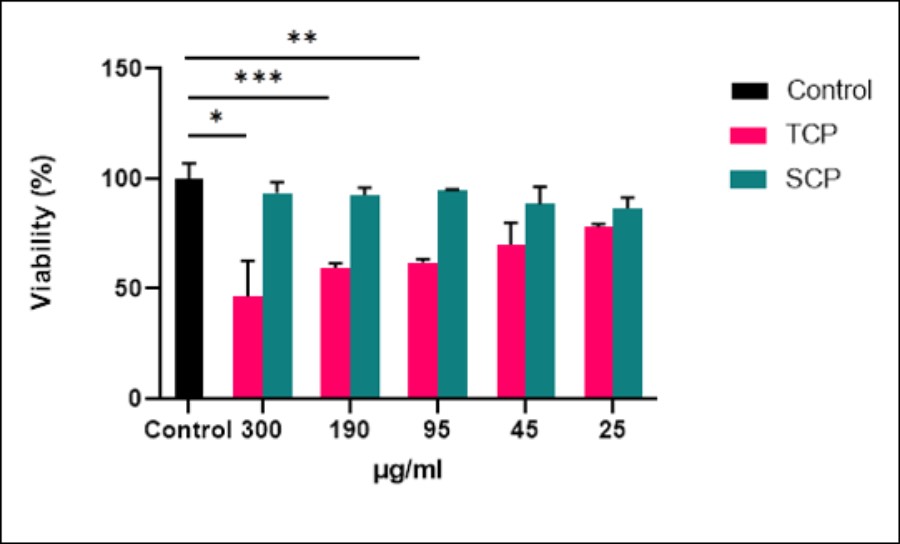 |
| Figure 6: MTT reduction viability test. Percentage of viability of mouse peritoneal macrophages (M?) purified from after incubation with different concentrations (300, 190, 95, 45 and 25μgml-1) of Total Cell Proteins (TCP) or Surface Cell Proteins (SCP). Results are expressed as the percentage of viability relative to untreated M? control. *, p=0.041 (300 μgml-1 of TCP), **, p=0.019 (190 and 95 μgml-1 of TCP), NS (45 and 25 μgml-1 of TCP). NS for all the SCP concentrations tested |
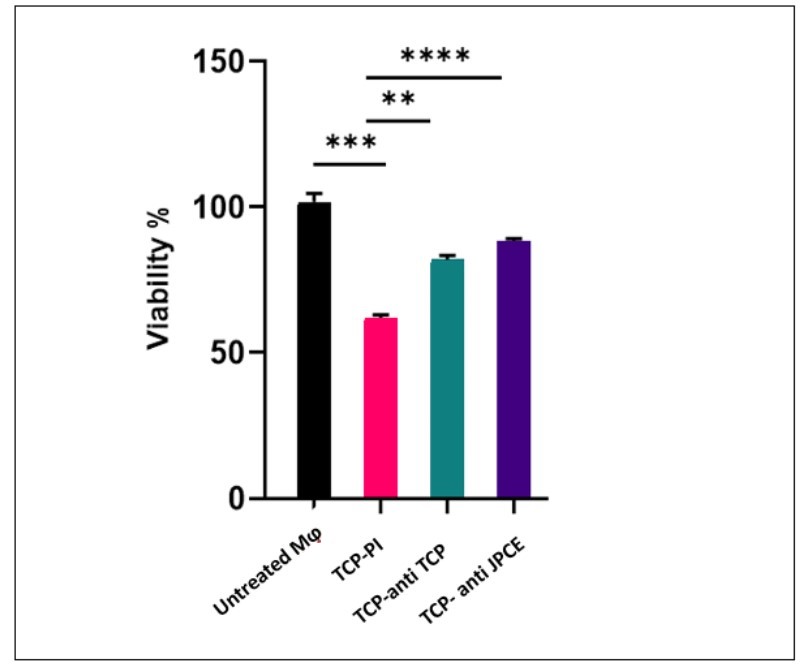 |
| Figure 7: MTT reduction viability test. Neutralization of C. albicans's Total Cell Protein (TCP) by anti-TCP and anti-JPCE sera. Percentage of viability of mouse peritoneal macrophages (M?) treated with 95 μgml-1 TCP pre-incubated with PI (preimmune sera), anti-TCP sera (1/50) and anti-JPCE sera (1/50). Results are shown as the percentage of relative viability with respect to control (untreated M?). **, p=0.006 (TCPPI), ***, p=0.0098 (TCP-anti-TCP sera), ***, p=0.0094 (TCP- anti-JPCE sera) |
|
Polyphagocytosis |
Associate Cell |
Phagocytosis |
Necrosis |
Anti–JPCE sera |
2.8 |
1.5 |
1.9 |
1.2 |
Anti-SCP sera |
||||
Pre-immune sera |




































