Open Access
Research Article
Max Screen
ISSN: 2638-082X
Copyright: © 2020 Bon LI. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Antenatal alcoholization leads to a number of specific disorders in the offspring organism, grouped into the concept of fetal alcohol syndrome, which is included in the fetal alcohol spectrum disorders. The negative effects of alcohol on the developing fetal brain include structural abnormalities of the brain, neurological, behavioral and cognitive defects. The cerebral cortex is especially sensitive to antenatal exposure to ethanol. It causes a decrease in the number and size of pyramidal neurons, a decrease in their protein content and underdevelopment of the cytoplasm, deep invaginations of the nuclear envelope, disintegration of the Golgi complex cisterns into vacuoles, swelling of mitochondria, an increase in the number of pathological forms of neuron.
The aim of this study was to estimate the ultrastructure of the internal pyramidal neurons of the frontal cortex of 5-day-old rats after antenatal alcoholization.
The studies were carried out on female outbred white rats with an initial weight of 230 ± 20 g and their offspring. The required sections of the brain cortex were cut and prepared for subsequent electron microscopic examination.
In neurons of 5-day-old rats who underwent prenatal alcoholization, an expansion of the perinuclear space and an eccentric arrangement of the nucleolus were revealed. The cytoplasm of some neurons is poor in organelles, and in the cytoplasm of other cells, the relative number of free ribosomes increases. Tubules of the granular endoplasmic reticulum often form an unusual network-like structure. Polymorphism of mitochondria and their hypertrophy were observed.
Antenatal alcoholization causes deep and varied ultrastructural changes in the pyramidal neurons of the frontal cortex of 5-day-old rat. The revealed changes in neurons of the cerebral cortex often underlie the known neurological and behavioral disorders in animals after antenatal alcoholization.
Keywords: Antenatal Alcoholization; Ultrastructural Changes; Neurons
List of abbreviations:GER: Granular Endoplasmic Reticulum
Antenatal alcoholization leads to a number of specific disorders in the offspring organism, grouped into the concept of fetal alcohol syndrome, which is included in the fetal alcohol spectrum disorders. The negative effects of alcohol on the developing fetal brain include structural abnormalities of the brain, neurological, behavioral and cognitive defects. The cerebral cortex is especially sensitive to antenatal exposure to ethanol. It causes a decrease in the number and size of pyramidal neurons, a decrease in their protein content and underdevelopment of the cytoplasm, deep invaginations of the nuclear envelope, disintegration of the Golgi complex cisterns into vacuoles, swelling of mitochondria, an increase in the number of pathological forms of neurons [1-11]. In our previous studies, alcohol consumption in rats during pregnancy caused a thickening of the cortex and swelling of pyramidal neurons of the cerebral cortex of their offspring on the 5th day of postnatal development, revealed by histological examination at the light-optical level [4,12]. At the same time, it was of interest to elucidate the features of the ultrastructure of these swollen neurons' disorders.
The aim of this study was to estimate the ultrastructure of the internal pyramidal neurons of the frontal cortex of 5-day-old rats after antenatal alcoholization.
The studies were carried out on female outbred white rats with an initial weight of 230 ± 20 g and their offspring. All experiments were carried out taking into account the rules of work with the use of experimental animals. This study was approved by the Biomedical Ethics Committee of the Grodno State Medical University (Protocol No. 1, dated 11.03.2014). The animals were fed the standard ration of the vivarium. Rats of the experimental group throughout pregnancy (from the day of detection of spermatozoa in vaginal smears to delivery) received a 15% ethanol solution as the only source of drinking, and animals of the control group received an equivalent volume of water. The animals were kept in the vivarium, in ventilated room at a temperature of 22 o C, at air humidity 50%, illumination not more than 25 luxin. In one cage, there were no more than five individuals. The ration of the animals included grains of cereals and legumes: oats, millet, barley, peas, and succulent feed: carrots, beets, cabbage. With three meals a day, in the morning, you could give a grain mixture and greens, in the afternoon - milk, in the evening - soft and juicy feed; they were provided with easy access to food and water. Protocols were reviewed and approved by the Ethical Committee of the Grodno State Medical University (protocol No 1, 14.04.2013).
The average alcohol consumption by pregnant females was 4 ± 2 g / kg / day [4,12]. The rat pups were slaughtered on the 5th day after birth. The location of the frontal cortex in histological preparations of the rat brain was determined using a stereotaxic atlas [13]. The required sections of the brain cortex were cut out with a blade and placed in 1% osmium fixative on Millonig's buffer (pH = 7.4) for 2 hours at a temperature of +4oC. Then they were washed in a mixture of Millonig's buffer (20 ml) and sucrose (900 mg) for 10 minutes, were dehydrated in alcohols of increasing concentration: 10 minutes in 50% alcohol, 10 minutes in 70% alcohol, two servings of 80% alcohol for 10 minutes, two servings of 96% alcohol for 10 minutes, 15 minutes in alcohol-acetone mixture (1:1), two servings of acetone for 15 minutes, 45 minutes in acetone-resin mixture (1:3), two servings of acetone-resin mixture (1:1) for 45 minutes, three servings of acetone-resin mixture (1:3) for 45 minutes, three servings of resin (araldite M + araldite H + dibutyl phthalate + DMR-30) for 60 minutes and embedded in this potting resin mixture.
Semi-thin sections (about 0.35 μm thick) were made using an MT-7000 ultramicrotome (RMC, United States), stained with methylene blue, and the sections of the inner pyramidal layer necessary for studying were cut out with a blade. Ultrathin sections (about 35 nm thick) were made on the same ultramicrotome, collected on supporting grids, and contrasted with uranium acetate and lead citrate. For this, the meshes with sections were dipped into a drop of uranyl acetate and kept for 20 minutes in the dark at room temperature, then washed in 3 portions of bidistilled water for 5 seconds and contrasted with lead citrate for 8 minutes, washed in 3 portions of bidistilled water, 5 seconds. The obtained preparations were studied in a JEM-1011 electron microscope (JEOL, Japan) and photographed with an Olympus MegaView III digital camera (Olympus Soft Imaging Solutions, Germany).
Each group had 5 rat pups, each animal. At least 20 neurons of the fifth layer of the cerebral cortex and the pyramidal layer were evaluated in each animal and 50 mitochondria were examined in each group, which ensured a sufficient sample size for subsequent analysis.
For statistical analysis of the data obtained in the experiment, non-parametric statistics methods were used (Statistica 10.0 software for Windows, StatSoft, Inc., USA). The results are presented in the form Me (LQ; UQ), where Me is the median, LQ is the boundary of the lower quartile; UQ is the boundary of the upper quartile. The differences between groups were considered significant at p< 0.05 (Kruskal-Wallis test with Bonferroni correction).
The average weight of the rat pups was 8 grams, the brain weight was 0.5 grams. There were no statistically significant differences between the control and experimental groups.
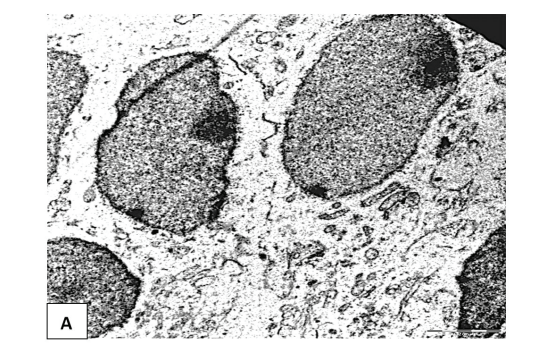
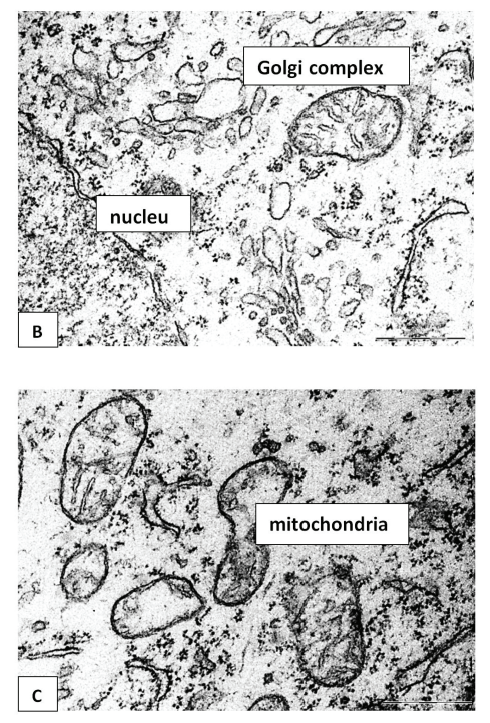
In 5-day-old control rats, normochromic neurons predominate in the inner pyramidal layer of the frontal cortex (Figure 1A). They are characterized by the presence of a large, euchromatin-rich cell nucleus with an eccentrically located nucleolus. Small clumps of heterochromatin are distributed evenly throughout the karyoplasm, an expansion of the perinuclear space was observed (Figure 1A). In the cytoplasm, tubules of the granular endoplasmic reticulum (GER) with a small number of ribosomes (Table 1), and a moderate number of mitochondria, whose cristae are not always well expressed, are detected. Dividing mitochondria were also identified (Figure 1B and C). The cisterns of the Golgi complex have not yet been formed at this stage of postnatal development. Free ribosomes in a moderate amount are diffusely distributed in the cytoplasm (Figure 1B and C).
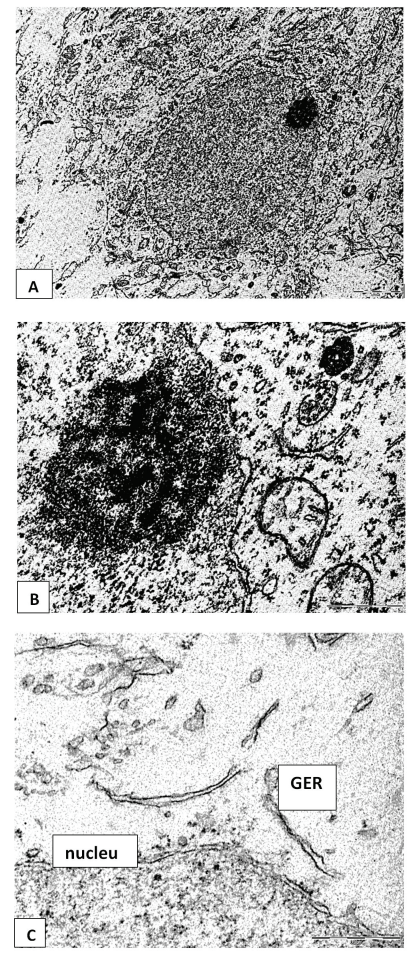
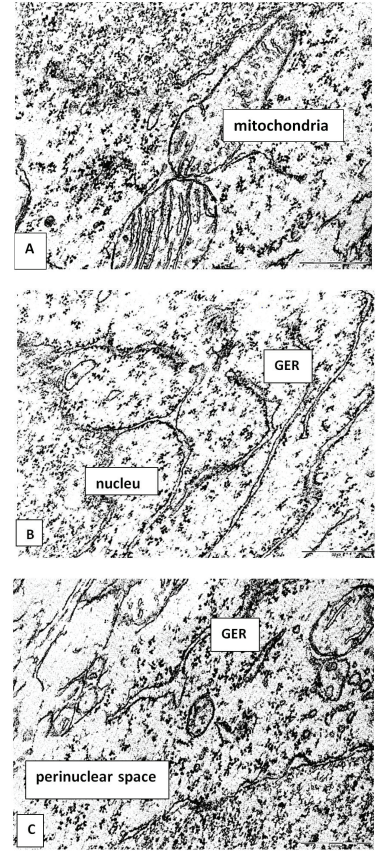
In neurons of 5-day-old rats who underwent prenatal alcoholization, an expansion of the perinuclear space and an eccentric arrangement of the nucleolus were revealed. The cytoplasm of some neurons is poor in organelles (Figure 2B), and in the cytoplasm of other cells, the relative amount of free ribosomes increases. GER tubules often form an unusual network-like structure; the lumen of their tubules sometimes narrows and widens unevenly. Polymorphism of mitochondria and their hypertrophy were observed (Figure 3)
In general, the neurons of the control animals had a normal ultrastructure typical of large pyramidal neurons of the rat cerebral cortex on the 5th day of postnatal development [14]. The expansion of the perinuclear space and the eccentric arrangement of the nucleolus may indicate an active metabolism of developing neurons in early ontogenesis.
In our preliminary studies in prenatally alcoholized animals 5 days after birth, a decrease in the total number of neurons was revealed at the light-optical level, as well as an increase in the size and swelling of preserved cells [4,12]. Disorganization of the endoplasmic reticulum cisterns, hyperplasia and their uneven expansion, as well as mitochondrial hypertrophy can be regarded as a manifestation of internal reparative processes.
Mitochondrial hypertrophy in the cytoplasm of neurons corresponds to the previously obtained histochemical data on the enzyme activation in them, which indicates an increase in the energy supply of cells [4,12].
A decrease in the number of ribosomes associated with GER and an increase in the number of free ribosomes indicate a switch in protein biosynthesis for the neurons' own needs, which is necessary for their survival in adverse conditions.
At the early stages of postnatal development, attempts to compensate for the negative processes of antenatal exposure to alcohol prevail. No gross violations of the ultrastructure of organelles were observed in 5-day-old rat pups. However, the data of our studies of the consequences of alcohol consumption during pregnancy showed that antenatal exposure to alcohol leads to significant qualitative and quantitative changes in the postnatal development of organelles of the internal pyramidal neurons of the frontal cortex of rats, compared with the control group. So, on the 20th, and especially on the 45th day, after birth, there is a decrease in the relative number of mitochondria, the number and length of their cristae, which indicates a violation of the energy supply of neurons. The relative number of free ribosomes in the cytoplasm of neurons in the cerebral cortex of antenatally alcoholized rats does not decrease with age, and the number of bound ribosomes does not increase. This indicates the predominance of protein biosynthesis for the own needs of neuronal perikaryons, to the detriment of protein biosynthesis for export, to their processes and terminals. At the same time, the number and size of lysosomes increase, which reflects an increase in autophagy processes in the neurons of prenatally alcoholized rats [9,15-17].
Thus, antenatal alcoholization causes deep and varied ultrastructural changes in the pyramidal neurons of the frontal cortex of 5-day-old rat. Moreover, these disorders look not only as a consequence of the direct damaging effect of alcohol, its metabolite acetaldehyde, or the oxidative stress induced by them on the membranes and organelles of neurons during embryogenesis, but also as a violation of a certain "program of development" of neurons of the cerebral cortex in postnatal ontogenesis, possibly through genetic and epigenetic mechanisms. The increased sensitivity of the developing brain to alcohol can be explained by the peculiarities of alcohol oxidation both in the mother's body during pregnancy and in the fetal brain. The revealed structural changes in pyramidal neurons of the cerebral cortex can underlie the known irreversible neurological and behavioral disorders in animals after antenatal alcoholization.
Funding: Grodno State Medical University grant for reagents and animals

![]()

  |
| Figure 1: Neurons of the inner pyramidal layer of the frontal cortex of control 5-day-old rats; (A) general view of the neuron; (B and C) Fragments of the nucleus and cytoplasm of the neuron; (C) Dividing mitochondria; Magnification: A - 8000, B, C - 50,000. Scale segment: A -2 μm, B, C - 0.5 μm. Electronograms |
 |
| Figure 2: Neurons of the inner pyramidal layer of the frontal cortex of 5-day-old rats subjected to antenatal alcoholization; (A) General view of the neuron; (B and C) Fragments of the nucleus and cytoplasm of the neuron; Magnification: A - 8000. B, C - 50,000. Scale segment: A -2 μm. B, C - 0.5 μm. Electronograms |
 |
| Figure 3: Neurons of the inner pyramidal layer of the frontal cortex of 5-day-old rats subjected to antenatal alcoholization; (A,B and C) Fragments of the nucleus and cytoplasm of the neuron; (B) GER exit from karyolemma, tubules of GER form an unusual network-like structure. Magnification: A, B, C - 50,000. Scale segment: A, B, C - 0.5 microns. Electronograms |
Indicators |
control |
alcohol |
Ribosome |
||
Total amount per 1 μm2 |
15,6 (14,6; 15,8) |
16,4 (16; 17,6)* |
Free per 1 μm2 |
13,8 (13,2; 14) |
14,9 (14,8; 16,2)* |
Related per 1 μm2 |
1,6 (1,4; 1,8) |
1,4(1,2; 1,4)* |
GER |
||
Tubules length, μm |
0,39 (0,38; 0,4) |
0,43 (0,42; 0,6)* |
Tubules width, μm |
0,035 (0,03; 0,04) |
0,04 (0,03; 0,05) |
Mitochondria |
||
Quantity in 1 μm2 |
0,8 (0,6; 0,8) |
0,8 (0,8; 1) |
Area, μm2 |
0,075 (0,073; 0,08) |
0,076 (0,07; 0,08) |
Form factor |
0,88 (0,87; 0,89) |
0,9 (0,889; 0,9) |
Elongation factor |
1,37 (1,26; 1,5) |
1,32 (1,3; 1,6) |
The number of cristae in mitochondria per 1 μm2 |
53 (41; 53) |
53 (27; 75) |
Total length of cristae in mitochondria per 1 μm2 |
8 (6; 8) |
8 (4; 11) |
The area occupied in the cytoplasm, μm2 |
0,06 (0,04; 0,064) |
0,06 (0,059; 0,08) |
Perinuclear space |
||
Size |
0,034 (0,03; 0,038) |
0,04 (0,04; 0,05)* |




































