Open Access
Research Article
Max Screen
ISSN: 2638-082X
Copyright: © 2020 Jing L. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Immunodepletion of high-abundant proteins (HAPs) aids in the identification and analysis of low-abundant proteins (LAPs) in complex samples. Currently, immunodepletion methods for bovine samples are very limited whereas greater availability exists for human and murine animal samples. In this study, we report the simultaneous depletion of HAPs from bovine samples using an immunoaffinity depletion cartridge (the multiple affinity removal system, MARS) designed to target six human HAPs, while such a depletion kit for bovine samples is not available. Sandwich ELISA analysis showed 98.65% to 99.98% depletion of albumin, IgG, and IgA from bovine plasma, follicular fluid (FF), ejaculated seminal plasma, epididymal fluid, and two mixtures of proteins (from washing ejaculated and epididymal spermatozoa with the high ionic strength solution). The immunodepletion method also dramatically depleted alpha1-antitrypsin, which is a target protein of the MARS technology, and also bovine binder of sperm proteins (BSPs) - PDC-109, BSP-A3, and BSP-30kDa. The BSPs are HAPs in ejaculated seminal plasma, forming 40%-57% of the total protein. The highest depletion rate among the BSPs was observed for the PDC-109 protein. The method has high reproducibility and can be incorporated into proteomic workflows to increase the sensitivity of proteomics analysis of LAPs in bovine biological fluids.
Keywords: Proteomics, High-Abundant Proteins (i.e., Albumin, IgG, IgA, and Alpha-1-Antitrypsin), Multiple Affinity Removal System (MARS), Bovine Fluids, Immunodepletion
Proteomics is the large-scale study of protein expression, structure, function and post-translational modifications in a cell or tissue. Advancements in proteomic sample preparation technologies, mass spectrometry instrumentation and bioinformatics [1-3] have increased the application of proteomics to understand various biological systems. Application of proteomics to bovine biological samples is an area of increasing interest [4]. Of particular interest is the application of proteomics to molecular research in cattle reproduction. This is important because reproductive performance (fertility) defines the attained biological and economic benefits. Potential benefits from such proteomic studies are extensive. Proteomics study can identify biomarkers for (i) detecting early pregnancy, and monitoring embryonic development and survival [5,6], (ii) evaluating quality of sperm, oocyte and cumulus cells [7-9], and (iii) characterizing the morphological changes during the pre-implantation period of pregnancy [10]. Results from the proteomic research can provide complementary information to traditional methods for assessing productivity in cattle.
The complexity and wide dynamic range in protein concentrations found in biological samples such as that from bovine present challenges in their preparation for proteomic analysis [11]. The wide dynamic range in protein concentrations results in highabundant proteins (HAPs, 1-100 mg/mL) masking the identification and characterization of low-abundant proteins (LAPs, < 100 ng/mL). However, biomarkers of disease and normal states are most probably of the low abundant types [12], which make the ability to characterize these LAPs imperative in proteomic analysis.
To bridge the wide concentration range and reduce sample complexity, proteomic workflows often incorporate fractionation method(s). Immunodepletion is a popular fractionation method due to its simplicity, specificity, and reliability. Immunodepletion method involves using antibodies to remove target HAPs, which enhances the subsequent detection of LAPS. The removal of HAPs allows for the loading of LAPs at higher concentrations into one-dimensional (1-D) or two-dimensional (2-D) gel electrophoresis and liquid chromatography (LC) to increase the sensitivity of proteomics analyses [13,14]. The reliability of the immunodepletion method has led to advancement in immunodepletion technologies including commercialized kits for human [13] and murine biofluids [15-17]. However, methods for immunodepletion of HAPs in samples from other species, including bovine, are very limited.
In the absence of available immunodepletion technology, proteomic analyses of bovine biological samples have used gel-based 2-D gel electrophoresis and modified forms of 2-DE such as 2-D fluorescence differential gel electrophoresis (2D-DIGE) followed by mass spectrometry analysis [5,18,19]. However, incorporating immunodepletion of HAPs will enhance the gel-based approach and also decrease the number of fractionation steps often employed in shotgun proteomic analysis. Faulkner et al. [20] presented separate depletion methods for albumin and IgG, the two top abundant proteins in bovine plasma. However, combining depletions, each targeting an abundant protein, increases analysis time and can result in sample loss and contamination.
The objective of this study was to develop an efficient and reproducible immunodepletion method for simultaneous removal of HAPs in bovine fluids. We employed the high capacity multiple affinity removal system (MARS) originally designed to target six HAPs - albumin, IgG, IgA, transferrin, alpha-1-antitrypsin, and haptoglobin (Hp) in human biological fluids. The six target HAPs represent 85%-90% of the total protein mass in human serum. The MARS technology has been successfully utilized in many proteomic analyses of human biofluids [21-26]. Because there is currently no available technology specific for simultaneous removal of several HAPs from bovine biological fluids, our application of the MARS technology presents a viable alternative for meeting this limitation. Removal of several HAPs will enable deeper probing of low copied proteins in bovine fluids.
Bio-SafeTM Coomassie G-250 stain, Tris/glycine/SDS buffer, 2-mercaptoethanol (β-ME), Laemmli sample buffer and 4-20% CriterionTM TGX precast gel were purchased from Bio-Rad Laboratories Inc. (California, USA). Formic acid, acetonitrile (ACN, LC/MS grade), 1 M triethylammonium bicarbonate (TEAB), and water (LC/MS grade) were obtained from Fisher Scientific (New Jersey, USA). Ammonium bicarbonate (ABC) and iodoacetamide were purchased from Acros Organics (New Jersey, USA). Dithiothreitol (DTT) and TPCK treated trypsin were purchased from Promega (Wisconsin, USA).
All procedures were approved by the South Dakota State University Institutional Animal Care and Use Committee. Samples in the present study were collected from a previous study to characterize changes in steroidogenic enzymes and follicular fluid (FF) steroid concentrations [27]. Briefly, 32 beef cows (i.e., cows raised for meat production) were synchronized by injecting with GnRH, 100 mg as 2 mL of Factrel, intramuscularly, Pfizer Animal Health (Madison, NJ, USA) on day -7 and prostaglandin F2α (PGF2α), 25 mg as 5 mL of Lutalyse intramuscularly (Zoetis, Florham Park, NJ, USA) on day 0. Estrus was monitored every 3 h from PGF2α on day 0 until hour 33 and at slaughter (hour 36 to 42) with the aid of EstroTect (Western Point, Inc, Apple Valley, MN, USA) estrus detection aids. Ovaries of all cows were examined on day -7, -4, and 0 by transrectal ultrasonography using an Aloka 500V ultrasound with a 7.5-MHz linear probe (Aloka, Wallingford, CT, USA) to assess follicular dynamics and ovulatory response. Ten cows that were determined to initiate a new follicular wave by day -4 were slaughtered on day 2 (hour 36 to 42) for ovary collection.
Immediately after ovary collection, follicular fluid (FF) was aspirated from dominant follicles (DF, >10mm diameter) and the granulosa cells (GCs) were separated from the FF by centrifugation (1,000 x g for 1 min). The FF was placed in RNase Free Tubes (USA Scientific), snap frozen in liquid nitrogen and stored at -80 o C until ready for analysis. Blood samples were collected at slaughter to provide better comparison to the FF collected at slaughter. To obtain plasma, blood collected at slaughter was placed in EDTA vacutainer tubes (Beckman Dickerson) and centrifuged at 1,200 x g for 30 min at 4 o C. The plasma supernatant was snap frozen in liquid nitrogen and stored at -80 o C until ready for further analysis.
All procedures were approved by the South Dakota State University Institutional Animal Care and Use Committee. Ejaculated semen from bulls (n = 9) were collected by electro-ejaculation weekly for three consecutive weeks. Collected ejaculated semen on the second week was centrifuged at 700 x g for 10 minutes to separate the spermatozoa from the seminal plasma. The seminal plasma fraction was labeled as “ejaculated semen plasma”. The remaining spermatozoa fraction was washed with a high ionic strength solution [28] and vortexed for one minute to remove any proteins attached and then centrifuged at 700 x g for 10 min. The supernatant was labeled as “ejaculated sperm proteins”. Following third sperm collection, bulls were rested for six weeks to allow epididymal reserves to renormalize. At the end of the resting period, bulls were slaughtered and the testes and epididymides were collected. Collected epididymides were dissected, and epididymal fluid and sperm were collected from the cauda section and processed as described for the ejaculated semen. The epididymal fluid fraction collected after separation from the epididymal spermatozoa was labeled as “epididymis semen proteins”. The spermatozoa were then washed as described above and the liquid fraction from washing off proteins on the epididymal spermatozoa was labeled as “epididymis sperm proteins”. All samples were stored at -80 oC.
Depletion of the different bovine fluids (PL, FF, epididymis sperm proteins, epididymis semen proteins, ejaculated sperm proteins, and ejaculated semen plasma) were performed using the High Capacity Multiple Affinity Spin Cartridge (MARS Hu-6HC) # 5188- 5341 (Agilent Technologies, CA, USA). The MARS Hu-6 system contains polyclonal antibodies designed to remove six HAPs - albumin, IgG, IgA, transferrin, alpha-1-antitrypsin, and Hp in human biological fluids. The depletion was done with vendorprovided buffers. Initial depletions used 2, 4, 6, 8, 10, 12, and 14 µL of bovine plasma and 4, 5, 7, 10, 12, and 14 µL of bovine FF to determine optimal volume for depletion. Each plasma and FF sample volume was diluted with 1x buffer A, pH 7.4 (# 5185-5987, Agilent Technologies, CA, USA) to achieve a final volume of 200 µL.
Initial quantification of the crude samples showed that ejaculated sperm proteins, ejaculated semen plasma, epididymis sperm proteins, and epididymis semen proteins were markedly of lower concentration compared to that of plasma or FF and upon depletion resulted in very low yield. Therefore, to ensure enough proteins were obtained in depleted samples for downstream analysis, depletion volumes were increased to 60 µL for ejaculated semen plasma and 154 µL each for ejaculated sperm proteins, epididymis semen proteins, and epididymis sperm proteins. For 154 µL depletion volume, 46 µL of 4x buffer A (# 5188-8283, Agilent Technologies, CA, USA) was added to achieve a total volume of 200 µL. For depletion volume of 60 µL, 94 µL of water was added after adding 46 µL of the 4x buffer A to obtain a final volume of 200 µL. The 4x dilution buffer A was used for the higher depletion volumes of these four samples to ensure a similar pH to the plasma and FF samples prepared using 1x buffer A. Diluted samples were transferred to 0.22 µm spin filters (Corning Incorporated, NY, USA) and centrifuged at 16,000 x g for one minute. Filtered samples were applied to the spin cartridge which was previously equilibrated with 4 mL 1x buffer A. Flow-through (FT) fraction was collected by centrifuging at 100 x g for 1.5 minutes. To ensure optimal recovery of FT, the cartridge was washed twice, each with 400 µL 1x buffer A. Each wash volume was collected by centrifuging for 2.5 minutes at 100 x g. Collected wash volumes were combined with initial collected FT. Bound fraction was eluted using 2 mL of elution buffer B (#5185-5988, Agilent Technologies, CA, USA). Non-depleted samples, bound fractions, FTs, and buffers were stored on ice throughout the experiment period to ensure protein integrity. Collected FTs and bound fractions were stored at -80oC until ready for analysis.
Coomassie (Bradford) Protein Assay Kit (#23200, Pierce Biotechnology, IL, USA) was used to determine protein concentrations of bound fractions, FT fractions, and non-depleted samples. The quantification was done according to manufacturer’s instruction. Bovine serum albumin, 2 mg/mL (#23209, Thermo Scientific, IL, USA), was used as a standard for making a calibration curve which covered a concentration range of 0.00-1500 µg/mL.
Concentrations of BSA, IgG, and IgA in non-depleted samples and FT fractions were determined using sandwich ELISA assay kits (Bethyl laboratories Inc., TX, USA). IgG was analyzed with bovine IgG ELISA quantitation set (Cat. No. E10-118), IgA with bovine IgA ELISA quantitation set (Cat. No. E10-131) and albumin with bovine albumin ELISA kit (Cat. No. E11-113). Samples were diluted and washed with vender-provided buffers. Dilutions were made based on protein concentration in samples. Specific dilution factors used for the different samples are shown in Table 1. All quantifications were performed according to manufacturer protocols. Briefly, 100 µL of dilute IgA or IgG Coating Antibody solution was added to each assay well. The wells were incubated at room temperature for an hour and washed with 1x Wash Buffer. 200 µL Blocking Solution was added to each well and incubated at room temperature for 30 minutes to block non-specific binding sites. Wells were then washed with 1x Wash Buffer. Assay wells for determining BSA concentration were pre-coated with anti-bovine albumin antibody. 100 µL of the diluted FT fractions or nondepleted samples were then added to the assay wells with the coated antibodies. After one hour incubation at room temperature, unbound proteins were washed with 1x Wash Buffer.
In determining IgA or IgG concentrations, 100 µL of horseradish peroxidase (HRP) conjugated anti-bovine IgA or IgG Detection Antibody solution was added to each well and incubated at room temperature for an hour to bind to the captured IgA or IgG. For albumin determination, 100 µL of Albumin Detection Antibody solution and 100 µL of HRP solution were added stepwise with incubations at room temperature for an hour after the Detection Antibody addition and for 30 minutes after the HRP solution addition. Wells were washed with 1x Wash Buffer after each step. 100 µl of 3,3’,5,5’-Tetramethylbenzidine (TMB) Substrate Solution was added to each well and incubated in darkness for 15 minutes in IgA or IgG measurements or 30 minutes in albumin measurement to initiate a calorimetric reaction, which was stopped by addition of 100 µL Stop Solution to each well. Absorbance was measured at 450 nm by Synergy H1 Hybrid Multi-Mode Reader (BioTek, VT, USA). Concentrations of BSA, IgA, and IgG were derived from calibration curves of reference standards included in the quantitation kits.
Flow-through and bound proteins were buffer exchanged into 0.05 M TEAB using Zeba Spin Desalting Columns, 7k MWCO (# 89891, Pierce Biotechnology, IL, USA) according to manufacturer protocol. Samples were vacuum dried prior to SDS-PAGE analysis. SDS-PAGE was performed under reducing and non-reducing conditions. Under reducing conditions, β-ME was diluted 1:20 in Laemmli buffer and the resulting solution was diluted twice with water. 15 µL of the prepared sample buffer was added to each vacuum dried sample. Samples were vortexed, centrifuged and then boiled in a water bath for three minutes. Boiled samples were quickly loaded onto a pre-cast gel. Non-reducing conditions excluded adding β-ME and heating steps. Equal amounts of 9 or 20 µg of proteins and 5-8 µL of Precision Plus ProteinTM unstained standard, 10-250 kDa (Bio-Rad Laboratories Inc., CA, USA) were used in the gel runs. Gels were electrophoresed in Bio-Rad PowerPacTM unit using 1x Tris/glycine buffer and at constant voltage of 150 for 62 minutes. Gel bands were visualized using Bio-SafeTM Coomassie G250 stain according to manufacturer instructions. Images were acquired using Proteineer SPII (Bruker Daltonics) equipped with SP3 Control software.
Identities of protein gel bands from bound fractions (HAP fractions) were determined using in-gel digestion and nano LC-MS/ MS analysis. For in-gel digestion, the gel band from the bound protein lane was excised with a sanitized cutting blade, transferred onto previously sanitized glass plate and then chopped into pieces. Gel pieces were transferred to a 0.5 mL microcentrifuge tube and washed with 100 µL LC/MS grade water with gently vortexing for five minutes. The water was removed and the gel pieces were completely destained by washing with several 100 µL portions of 25 mM ammonium bicarbonate (ABC) in ACN/water (v/v = 50/50) solution. Destained gel pieces were dehydrated with 100 µl ACN for 10 minutes and dried in a speed-vac at 4oC for 5 minutes. Reduction was done with 100 µl of 10 mM DTT at 55oC for 30 minutes.
After removal of reduction solution, alkylation was carried out by adding 100 µL of freshly prepared 55 mM iodoacetamide and incubating in a dark place for 45 minutes. Gel pieces were washed with 100 µL 25 mM ABC solution for 10 minutes, dehydrated with 100 µL ACN, and dried in speed-vac at 4oC for five minutes. 30 µL of freshly prepared cold 20 µg/mL TPCK treated trypsin (P/N 4370282, AB Sciex Pte Ltd, MA, USA) in 25 mM ABC solution was then added to each sample and incubated on ice until gel pieces became fully rehydrated and clear. Excess trypsin solution was removed and the gel pieces were covered with 30 µL 25 mM ABC solution and incubated for 16 h at 37oC. To extract tryptic peptides, 25 µL of 25 mM ABC solution was added and the liquid fraction was collected. 50 µL of ACN was then added and incubated for 10 minutes with slightly shaking and the liquid fraction was also collected. Finally, 30 µL of water and 50 µL of ACN were added stepwise with 10 minutes of incubation with shaking in each step, and the liquid fraction in each step was also collected. All extracted liquid fractions were combined, frozen for 15 minutes and dried in a speed-vac at 4oC. Dried peptides were stored at -80oC.
Dried peptides were purified and concentrated with either PierceTM C18 spin columns (Pierce Biotechnology, IL, USA) or ZipTip C18 tips (Millipore, MA, USA) based on high intensity or low intensity of Coomassie stained gel bands. Purification was done according to manufacturer protocols. Purified samples were dried in a speed-vac at 4oC and stored at -80oC until analysis. Protein identification was performed with nano-liquid chromatography tandem mass spectrometry (nanoLC-MS/MS) analysis by using Eksigent nanoLC - Thermo Fisher LTQ mass spectrometer. The dried, purified digest from each interested protein band was brought up in water/ACN/formic acid (95%/5%/0.1%) and then loaded on IntegraFrit sample trap (ProteoPepTM II C18,300 Å, 5 µm, 100 μm × 25 mm, New Objective, Inc., Woburn, MA). The retained peptides were washed isocratically with water premixed with 0.1% formic acid pumped from channel 1A to remove any excess reagents. The cleaned peptides were resolved on an IntegraFrit analytical column (ProteoPepTM II C18, 300 Å, 5 µm, 75 μm ×100 mm, New Objective, Inc., Woburn, MA) with a multistep gradient of solvent 2A (water premixed with 0.1% formic acid) and solvent 2B (ACN premixed with 0.1% formic acid) at a flow rate of 200 nL/min. The LTQ mass spectrometer was operated in the data-dependent mode. The full MS spectra were acquired in positive mode with mass range of 300-1800 m/z. Top three ions with intensities exceeding a preset threshold in full mass scan were chosen for the followed MS/MS scan.
The LC-MS/MS raw data from nanoLC-LTQ were converted to DTA files using Thermo Proteome Discoverer and correlated to theoretical fragmentation patterns of tryptic peptide sequences from the FASTA database (bovine) using SEQUESTTM (Thermo Fisher). The search parameters included: (i) fixed cysteine modifications of +57 Da for carbamidomethyl-cysteines, (ii) variable modifications allowing +16 Da with methionines for methionine sulfoxide, (iii) restricted to trypsin-digested peptides and allowed for two missed cleavages, (iv) precursor mass tolerance of 2.5 Da and fragment mass tolerance of ±0.8 Da, and (v) target false discovery rate (FDR) strict of 0.01 and FDR relaxed of 0.05. Peptides were identified with FDR below 0.05 and individual cross correlation exceeding a threshold dependent on the precursor charge state. The proteins matched with at least two peptides were considered as positive identification. Some proteins identified from FASTA database (bovine) were uncharacterized. These uncharacterized protein’s names/families were derived with UniProtKB by comparing to human versions of the bovine proteins using accession numbers.
To determine reproducibility of the MARS Hu-6HC cartridge, the bovine fluids were subjected to 3-5 replicate depletions on same day for 2-5 separate days. Reproducibility was assessed by the coefficient of variations (CVs) of the percentage of total protein depleted and by SDS PAGE separation of crude, bound and FT fractions.
Percent depletion of the bovine samples by using the MARS Hu-6HC cartridge in removing the target HAPs was determined by comparing the total protein content in the FT fraction to that of the non-depleted samples. The percentage of total protein depleted from each bovine fluid is shown in Table 2. Total proteins depleted, expressed as percent depletion, were comparable for different volumes of plasma or FF investigated: 67.9 ± 3.1% (4.1%) for 2, 4, 6, 8, 10, 12, and 14 µL of plasma and 83.4 ± 6.0% (7.2%) for 4, 5, 7, 10, 12, and 14 µL of FF; mean ± SD (%CV) (Figure 1). Therefore, 4 µL of plasma and 5 µL of FF were used in further investigation to conserve samples. The amount of proteins and LAPs (yield) in the load volume of each bovine fluid, and the concentration and percent of LAPs (yield) in each bovine fluid are shown in Table 3. From two to five separate days of depletion, percentage of total proteins depleted follows the order FF (82.6 ± 3.8%) > plasma (72.1 ± 3.8%) > epididymal sperm proteins (66.1 ± 4.6%) > epididymis semen proteins (61.3 ± 3.8%) > ejaculated semen plasma (53.7 ± 1.9%) > ejaculated sperm proteins (49.8 ± 3.3%) (Table 2). Similar values were obtained for same day depletions (Table 2).
The effectiveness of depletion of abundant proteins was also assessed by SDS PAGE separation of non-depleted samples, bound and FT fractions. Figures 2 and 3 reveal the unmasking and highlighting of LAPs after dramatic removal of HAPs from the different bovine fluids. NanoLC-MS/MS analysis of gel bands from bound fractions identified four out of the six target HAPs, namely serum bovine albumin (BSA), IgG, IgA and α-1-antitrypsin. Gel bands containing these proteins are shown in Figure 2. Transferrin and Hp were not identified in the bound fractions by the nanoLC-MS/MS method.
Depletion efficiency (i.e., the extent of removal) of BSA, IgG, and IgA from the different bovine fluids was individually assessed using sandwich ELISA. Results showed depletion of 98.73% to 99.98% BSA, 98.65% to 99.96% IgG, and 98.79% to 99.96% IgA from all the bovine samples studied (Table 4). The observed depletion rates are comparable to manufacturer specification of 98.9%- 99.9% obtained for human biological fluids. Depletion efficiency for alpha-1-antitrypsin was not measured by the ELISA method.
To assess the specificity of the depletion method for the target proteins in bovine fluids, 1-D SDS-PAGE and nanoLC-MS/MS analysis were performed on the bound fractions. Concomitant removal of non-target proteins was observed from all the bovine samples depleted. These are summarized in Table 5. Non-target proteins from the bound fractions of the ejaculated sperm proteins and ejaculated semen plasma included three major binder of sperm proteins (BSPs), namely PDC-109, BSP-A3, and BSP-30kDa (Figure 3). Among the BSPs depleted, the highest depletion rate was observed for PDC-109, which forms 25%-47% of seminal plasma total proteins [29].
The immunodepletion method had good reproducibility for all the bovine samples studied. The coefficient of variations (%CVs) of depletion efficiencies determined on the same day and separate days ranged from 3.2% to 6.8% and 3.5% to 6.9 %, respectively (Table 2). The good reproducibility was also supported by the consistent protein band pattern on the SDS PAGE gel.
Immunodepletion methods are popular and often the first step in fractionation to narrow the wide concentration range in biological samples and reduce sample complexity. To date, available immunodepletion methods for bovine samples are very limited. This study reports a method for simultaneous immunodepletion of abundant proteins BSA, IgG, IgA, and alpha-1-antitrypsin from bovine plasma, FF, epididymis sperm proteins, epididymis semen proteins, ejaculated semen plasma, and ejaculated sperm proteins with high depletion rates. Depletion rates for BSA, IgG, and IgA were confirmed as 98.65% to 99.98% from all the bovine fluids studied using sandwich ELISA. The observed depletion rates are comparable to the 98.9%-99.9% depletion rates for these proteins from human fluids as stated by the manufacturer. The similar depletion rates indicate that sample variability does not significantly affect the affinity of the MARS polyclonal antibodies for the target proteins in bovine fluids. Sequence similarities between the target proteins in human and bovine explain the effective application of this immunodepletion technology on bovine fluids. A search in UniprotKB database (http://www.uniprot.org) showed that human serum albumin and alpha-1-antitrypsin respectively share ≥75% and ≥68% sequence identity with their respective proteins in bovine. Sequence identity between human and bovine immunoglobulin light chains range is 53%-76%, and the heavy chains range is 33%-65%. The high depletion rates achieved with the presented method make it a viable alternative for achieving immunodepletion of these abundant proteins in bovine biological fluids while similar technology is currently unavailable.
The total proteins depleted from a sample seem to depend on the source of the proteins, i.e., in human or cattle. Total proteins depleted were about 50%-83% for the bovine samples investigated. This range was lower than the manufacturer specification of 85%-90% for human serum, but total proteins depleted from bovine FF (82.6%) and plasma (72.1%) were closer to the stated lower end range. Total proteins depleted were based on the levels of the target proteins in certain samples. Thus, differences in the levels of the target proteins in the different bovine fluids and between human and bovine would affect the total proteins depleted. For instance, albumin, the most abundant protein in plasma represents 55% of the total proteins in human plasma [12] and about 42% in bovine plasma [30]. Plasma and FF are closely related in composition and majority of proteins in FF are viewed to originate from the blood [31-33]. However, albumin concentration is higher in FF compared to serum [34,35], which might explain the high depletion rate attained with the FF. 2-DE analysis of caudal epididymis fluid found albumin represented ~21% of all proteins spots detected [36]. By contrast, albumin concentration is very minimal in seminal plasma [37]. Accordingly, total proteins depleted were lower for the ejaculated semen plasma sample compared to the plasma and FF samples. Total proteins depleted were lower for ejaculated seminal plasma compared to epididymal fluid (named epididymis semen proteins).
The lower total proteins depleted from bovine fluids are also attributable to the lack of depletion of Hp and transferrin. Both are target proteins of the MARS Hu-6HC cartridge. Sequence homology could not explain the lack of removal of both proteins as they are highly homologous between human and bovine, 69% and 75% sequence identity, respectively. High specificity of the MARS Hu-6HC cartridge for human transferrin and Hp appears to be the primary reason for both proteins not being depleted from the bovine fluids. The lack of depletion of Hp can also be influenced by the very low level of this protein under non-inflammatory conditions (< 0.2 g/L) in cattle [38-40]. Although Hp and transferrin are not depleted, the simultaneous rapid depletion of albumin, IgG, IgA, and alpha-1-antitrypsin from the bovine fluids will facilitate the application of the current immunodepletion method to bovine proteomics. Albumin and IgG represent about 60% of the bovine plasma proteome. Depletion of both proteins increased the number of spots and intensities on a 2-DE map [20]. The additional depletion of abundant proteins by the presented depletion method is expected to provide more desirable probing of low copied proteins in gel-based and gel-free proteomic workflows for bovine fluid analysis. As shown in this study, the depletion of the four major proteins unmasked and highlighted more protein gel bands in SDS PAGE gels. The HAPs were removed in a single step, saving time and reducing sample loss.
To determine reproducibility of the immunodepletion, replicate depletions on same day and on separate days were analyzed. The results show that the method has high reproducibility on bovine fluids as revealed by the low % CV values of total proteins depleted from different and same volumes. Because immunodepletion is often the first step fractionation option, a good reproducibility is critical to the accuracy and further reproducibility of downstream protein separation, identification, and quantification. Low loading volumes of plasma and FF (4 µL and 5 µL) did not affect the reproducibility. This can be beneficial if the sample is limiting. The presented concentration of LAPs and amount of LAPs in such low load volumes may guide downstream experiments.
Concomitant removal of non-target proteins in immunodepletion methods is often inevitable [23,24]. Non-target proteins removal from the bovine fluids included other HAPs, e.g., PDC-109, BSP-A3, BSP-30kDa, alpha-2-macroglobulin, complement C3, complement C4, and fibrinogen. Non-target proteins’ removal could result from non-specific binding to cartridge resin, accidental/specific cross reaction of the antibodies, or interactions with the target proteins. BSPs were consistently removed from the ejaculated serum proteins and ejaculated semen plasma. Their dramatic removal is considered beneficial since they are major proteins constituting 40%-57% of the total protein in the seminal plasma component of ejaculates. PDC-109 alone forms 25%-47% of seminal plasma total proteins [29]. Thus, depletion of the BSPs can additionally enhance the analysis of LAPs in these ejaculates. However, caution should be taken when applying this depletion method to target these seminal proteins since their removal is non-specific. In such cases, a better approach would be to analyze both the bound and FT fractions to ensure adequate estimation of their depletions.
Immunodepletion methods for bovine fluids are currently very limited. This study describes an immunodepletion method for simultaneous removal of four HAPs (i.e., albumin, IgG, IgA, and alpha-1-antitrypsin) in different bovine fluids. The high depletion efficiencies (98.7%-99.9%) and reproducibility make it practical for its application in bovine proteomic analyses.
We thank the support of the National Science Foundation (NSF) grant number 1446886, and USDA/National Institute of Food and Agriculture, Hatch project 1004566. We also thank the Core Campus Mass Spectrometry Facility at South Dakota State University (SDSU) supported in part by the NSF/EPSCoR Grant No. 0091948 and NSF MRI Grant No. SS1000060 and the State of South Dakota. Finally, we thank the Functional Genomics Core Facility at SDSU supported in part by NSF/EPSCoR Grant No. 0091948 and by the State of South Dakota.
The authors declare no conflicts of interest related to this work or publication. There was no writing assistance in the production of this manuscript.

![]()

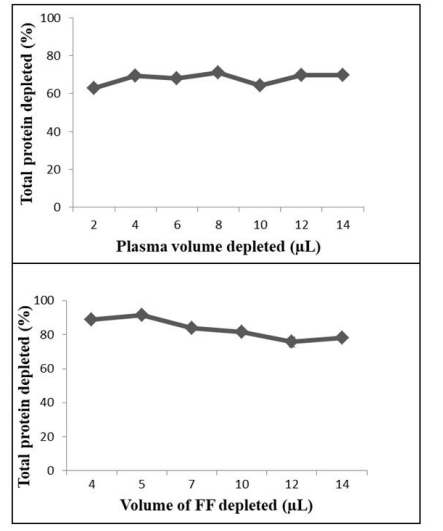 |
| Figure 1: Total proteins depleted from different volumes of bovine plasma and follicular fluid (FF) with the MARS Hu-6HC cartridge. Top figure shows that total proteins depleted are similar over 2 (n = 7), 4 (n = 7), 6 (n = 7), 8 (n = 2), 10 (n = 5), 12 (n = 2), and 14 (n = 8) μL of plasma. Bottom figure shows that total proteins depleted are similar over 4 (n = 1), 5 (n = 1), 7 (n = 3), 10 (n = 2), 12 (n = 2), and 14 (n = 2) μL of FF. |
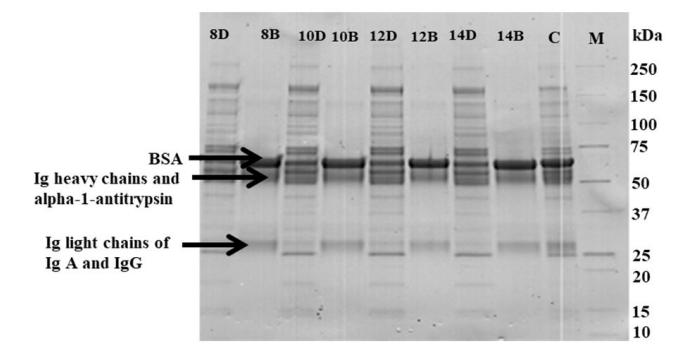 |
| Figure 2: Reproducibility of the MARS Hu-6HC cartridge on different depletion volumes of bovine plasma. Lanes: 8D, 10D, 12D, and 14D- Flow-through fractions from depletion of 8, 10, 12, and 14 μL of bovine plasma, respectively; 8B, 10B, 12B, and 14B- Bound fractions from depletion of 8, 10, 12, and 14 μL of bovine plasma, respectively; C- Crude bovine plasma; M- Molecular weight marker. Similar depletion efficiencies were obtained for the different volumes of plasma depleted. Arrows show gel bands containing BSA, IgA, IgG, and alpha-1- antitrypsin. 9 μg of proteins were loaded on each lane of 4-20% CriterionTM TGX precast gel and run under reducing conditions using β-ME. Bands were visualized with Coomassie blue staining. |
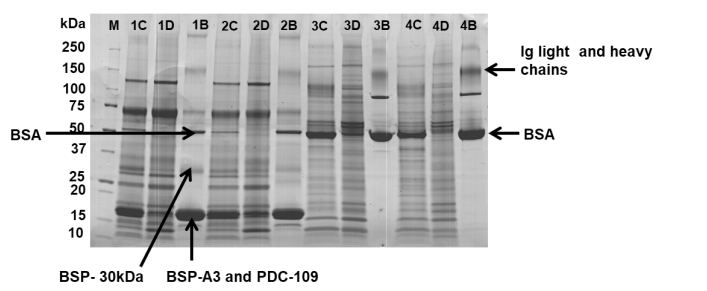 |
| Figure 3: Unmasking and highlighting of low-abundant proteins (LAPs) after removal of high-abundant proteins (HAPs) from bovine ejaculated and epididymis sperm proteins, ejaculated semen plasma, and epididymis semen proteins with MARS Hu-6HC cartridge. MMolecular weight marker; C- Crude sperm sample; D- Flow-through fraction; B- Bound fraction. 1, 2, 3, and 4 refer to ejaculated semen plasma, ejaculated sperm proteins, epididymis semen proteins, and epididymis sperm proteins, respectively. BSA and Ig were targeted proteins depleted. Binder of sperm proteins (BSPs) - PDC-109, BSP-A3, and BSAP-30 kDa were non-specifically depleted from ejaculated sperm proteins and seminal plasma. BSPs are HAPs found in these two samples. 20 μg of proteins were loaded on each lane of 4-20% CriterionTM TGX precast gel and run under non-reducing conditions. Bands were visualized with Coomassie blue staining. |
|
Dilution factors |
|||||
|
Albumin |
IgG |
IgA |
|||
Sample |
Non-depleted |
FT |
Non-depleted |
FT |
Non-depleted |
FT |
Plasma |
2.5E+05 |
4.0E+02 |
1.0E+05 |
2.0 |
7.5E+04 |
3.0 |
Follicular fluid |
1.5E+05 |
2.0E+01 |
9.0E+04 |
2.0 |
1.5E+04 |
2.0 |
Epididymis sperm proteins |
3.0E+03 |
1.0E+02 |
1.0E+02 |
2.0 |
5.0E+01 |
1.0 |
Epididymis semen proteins |
1.0E+04 |
3.0E+02 |
3.0E+02 |
8.0 |
5.0E+01 |
2.0 |
Ejaculated sperm proteins |
3.0E+03 |
5.0E+02 |
2.0E+02 |
2.0 |
1.0E+02 |
5.0 |
Ejaculated semen plasma |
1.0E+04 |
2.0E+01 |
3.0E+02 |
2.0 |
2.0E+02 |
1.5 |
Sample |
Volume depleted (µL) |
Total protein depleted on same day (%)a |
Total protein depleted on separate days (%)b |
||
|
|
Mean ± SD |
%CV |
Mean ± SD |
%CV |
Plasma |
4 |
71.9 ± 4.9 |
6.8 |
72.1 ± 3.8 |
5.2 |
Follicular fluid |
5 |
86.0 ± 3.6 |
4.2 |
82.6 ± 3.8 |
4.6 |
Epididymis sperm proteins |
154 |
66.2 ± 2.1 |
3.2 |
66. 1 ± 4.6 |
6.9 |
Epididymis semen proteins |
154 |
58.2 ± 2.9 |
5 |
61.3 ± 3.8 |
6.2 |
Ejaculated sperm proteins |
154 |
49.7 ± 2.9 |
5.9 |
49.8 ± 3.3 |
6.6 |
Ejaculated semen plasma |
60 |
53.6 ± 2.4 |
4.5 |
53.7 ± 1.9 |
3.5 |
Sample |
Crude (µg/ml)* Mean ± STD |
Load volume (µL) |
Load (µg)* Mean ± STD |
Yield (µg)* Mean ± STD |
Yield (%)* Mean ± STD |
Yield (µg/mL)* Mean ± STD |
Plasma |
63960.2 ± 6523.0 |
4 |
255.8 ± 26.1 |
71.2 ± 11.4 |
27.9 ± 3.8 |
17798. ± 2839.4 |
Follicular fluid |
56445 ± 4196.8 |
5 |
287.9 ± 19.9 |
47.9 ± 11.1 |
17.3 ± 4.2 |
9988.6 ± 2119.4 |
Epididymis sperm proteins |
582.8 ± 64.3 |
154 |
89.8 ± 9.9 |
30.5 ± 4.5 |
34.1 ± 4.6 |
200.5 ± 30.8 |
Epididymis semen proteins |
1857.6 ± 236.7 |
154 |
286.1 ± 36.5 |
109.7 ± 7.33 |
38.7 ± 3.82 |
712.6 ± 47.6 |
Ejaculated sperm proteins |
1744.6 ± 42.35 |
154 |
268.7 ± 6.52 |
134.6 ± 9.4 |
50.9 ± 3.4 |
874.2 ± 61.13 |
Ejaculated semen plasma |
19678.9 ± 2866.7 |
60 |
1238.2 ± 156.8 |
552.2 ± 66.4 |
44.6 ± 1.5 |
9202.9 ± 1106.9 |
|
% Depletion efficiency (Mean ± SD)* |
||
Sample |
Albumin |
IgG |
IgA |
Plasma |
98.73 ± 0.29 |
99.45 ± 0.01 |
98.85 ± 0.21 |
Follicular fluid |
99.97 ± 0.02 |
99.96 ± 0.00 |
98.85 ± 0.60 |
Epididymis sperm proteins |
99.92 ± 0.04 |
99.61 ± 0.24 |
98.79 ± 0.45 |
Epididymis semen plasma |
99.98 ± 0.00 |
99.57 ± 0.21 |
99.40 ± 0.12 |
Ejaculated sperm proteins |
99.52 ± 0.20 |
99.52 ± 0.03 |
99.63 ± 0.04 |
Ejaculated semen plasma |
99.45 ± 0.01 |
98.65 ± 0.06 |
99.96 ± 0.02 |
Accession |
No of peptides identified |
Score |
Description |
Protein familyᵻ |
B8Y9S9 |
38 |
528.63 |
Embryo-specific fibronectin 1 transcript variant |
|
P07589 |
27 |
515.40 |
Fibronectin |
|
Q7SIH1 |
33 |
395.84 |
Alpha-2-macroglobulin |
|
P02784a |
8 |
323.32 |
Seminal plasma protein PDC-109 |
|
F1MAV0 |
19 |
238.79 |
Fibrinogen beta chain |
|
A5PJE3 |
16 |
229.46 |
Fibrinogen alpha chain |
|
G3X7A5 |
17 |
179.49 |
Complement C3 |
|
P12799 |
8 |
173.91 |
Fibrinogen gamma-B chain |
|
P81019a |
3 |
62.58 |
Seminal plasma protein BSP- 30 kDa |
|
P01030 |
2 |
42.92 |
Complement C4 |
|
P04557a |
2 |
34.56 |
Seminal plasma protein A3 |
|
F1MBL6 |
2 |
32.17 |
Uncharacterized protein (Fragment) |
P13/P14 kinase |
F1MI18 |
2 |
24.05 |
Uncharacterized protein |
Alpha-2 macroglobin |
E1BIF6 |
2 |
19.75 |
Uncharacterized protein |
Ubiquitin associated domain |
P23805 |
4 |
16.95 |
Conglutinin |
|
Q28085 |
2 |
15.71 |
Complement factor H |
|
G3N0S9 |
2 |
11.42 |
Uncharacterized protein |
Sushi CCP/SCR domain |
F1N5M2 |
2 |
8.76 |
Vitamin D-binding protein |
|




































